

2 курс / Гистология / Общий курс гистологии
.pdfМикроглия представляет собой совокупность мелких удлиненных звездчатых клеток (микроглиоцитов) с плотной цитоплазмой и короткими ветвящимися отростками, располагающихся вдоль капилляров ЦНС. Они имеют мезенхимное происхождение, развиваются непосредственно из моноцитов и относятся к макрофагальномоноцитарной системе. Для них характерны ядра с преобладанием гетерохроматина и высокое содержание лизосом в цитоплазме.
Функция микроглии – защитная, в том числе иммунная. Микроглиальные клетки рассматривают как специализированные макрофаги ЦНС. Они обладают подвижностью, активируясь и увеличиваясь в числе при воспалительных и дегенеративных заболеваниях нервной системы. Клетки микроглии способны выделять цитокины и участвовать в иммунных реакциях.
Нервные волокна представляют собой отростки нейронов, покрытые глиальными оболочками. Различают безмиелиновые и миелиновые волокна, состоящие из центрально расположенного отростка нейрона (осевого цилиндра), окруженного оболочкой из клеток олигодендроглии (рис. 60). В периферической нервной системе они называются леммоцитами, или шванновскими клетками.
Безмиелиновые нервные волокна характеризуются низкой ско-
ростью проведения нервного импульса (0,5–2 м/с). Они образуются путем погружения осевого цилиндра (аксона) в цитоплазму леммоцитов, располагающихся в виде тяжей. Плазмолемма при этом прогибается, окружая аксон, и образует дупликатуру – мезаксон. Нередко в цитоплазме одного леммоцита могут находиться до 10–20 осевых цилиндров. Такое волокно называется волокном кабельного типа. Поверхность волокна покрыта базальной мембраной.
Рис. 60. Миелиновые и безмиелиновые волокна (по В.Л. Быкову).
1–3 – стадии формирования миелинового волокна. 4 – безмиелиновое волокно (кабельного типа). ЛЦ – леммоцит, МА – мезаксон, А – аксон, Я – ядро леммоцита, МО – миелиновая оболочка
161
Миелиновые нервные волокна встречаются в ЦНС и ПНС и характеризуются высокой скоростью проведения нервного импульса (5–120 м/с). Миелиновые волокна толще безмиелиновых и содержат осевые цилиндры большего диаметра. В миелиновом волокне осевой цилиндр непосредственно окружен миелиновой оболочкой, вокруг которой располагаются тонкий слой цитоплазмы и ядро леммоцита – нейролемма. Снаружи волокно покрыто базальной мембраной. Миелиновая оболочка возникает в результате слияния многочисленных мембранных витков (пластин). В состав миелина входят липиды в высокой концентрации.
Образование миелиновой оболочки в ПНС: погружение осевого цилиндра в леммоцит сопровождается формированием длинного мезаксона, который начинает вращаться вокруг аксона, образуя первые рыхло расположенные витки миелиновой оболочки. По мере увеличения витков в процессе созревания миелина они располагаются все более плотно и частично сливаются. Промежутки между ними, заполненные цитоплазмой леммоцита, сохраняются лишь в отдельных местах, не окрашиваемых осмием, – миелиновых насечках (Шмид- та-Лантермана).
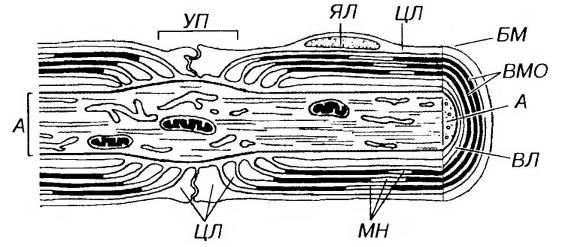
Перехваты Ранвье – участки в области границы соседних леммоцитов, миелиновая оболочка здесь отсутствует. Перехваты повторяются по ходу миелинового волокна через 1–2 мм. В области перехвата на плазмолемме присутствуют многочисленные натриевые каналы, которых нет вне перехватов (рис. 61).
Рис. 61. Ультраструктурная организация миелинового волокна (по В.Л. Быкову). А – аксон, УП – узловой перехват Ранвье, ЯЛ – ядро леммоцита, ЦЛ – цитоплазма леммоцита, БМ – базальная мембрана нервного волокна, ВМО – витки миелиновой оболочки, ВЛ – внутренний листок, МН – миелиновые насечки Шмидта-
Лантермана
162
Образование миелиновой оболочки в ЦНС: осевой цилиндр не по-
гружается в цитоплазму олигодендроцита, а охватывает его плоским отростком, который в дальнейшем вращается вокруг него, теряя цитоплазму, причем его витки превращаются в пластинки миелиновой оболочки. В отличие от шванновских клеток, один олигодендроцит может участвовать в миелинизации многих (до 40–50) нервных волокон.
Нарушение образования и повреждение образованного миелина лежат в основе ряда тяжелых заболеваний нервной системы. Миелин
вЦНС может повреждаться при аутоиммунных заболеваниях с его разрушением (демиелинизация). Этот процесс активно протекает при рассеянном склерозе – тяжелом заболевании, связанном с расстройством различных функций, развитием параличей, потерей чувствительности. При некоторых метаболических расстройствах возникают нарушения образования миелина – лейкодистрофии, проявляющиеся
вдетстве тяжелыми поражениями нервной системы.
Нервные окончания
Различают 3 группы нервных окончаний:
1)рецепторные (афферентные, или чувствительные), образованные дендритами;
2)эффекторные окончания (эфферентные, или двигательные), по которым нервный импульс передается на рабочий орган;
3)синапсы – места контактов нейронов, через которые осуществляется передача нервного импульса.
1. Чувствительные нервные окончания
Выделяют экстра- и интрарецепторы, соответственно воспринимающие информацию от покровных тканей и от внутренних органов. В зависимости от раздражителя выделяют механорецепторы, хеморецепторы, терморецепторы и осморецепторы, хотя строгой зависимости строения рецептора от действующего стимула не наблюдается.
Чувствительные нервные окончания подразделяют на свободные,
несвободные и инкапсулированные (рис. 62).
Выделение свободных и несвободных окончаний весьма условно и основано на данных Б.И. Лаврентьева, определившего свободные окончания как такие аппараты, где осевые цилиндры и их ветвления лежат свободно среди клеток эпителия или в промежуточном веществе соединительной ткани. Несвободные окончания окружены сравнительно небольшим количеством вспомогательных – «специальных»
– клеток, обеспечивающих трансформацию внешних раздражений в нервные импульсы.
163
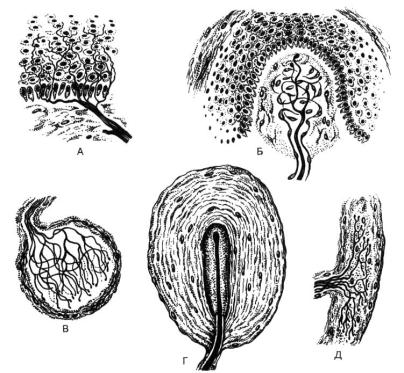
Рис.62. Рецепторные нервные окончания (по Ю.И. Афанасьеву).
А – свободные нервные окончания (боль), Б – тельце Мейснера (прикосновение), В – колба Краузе (холод), Г – пластинчатое тельце Фатера-Пачини (давление), Д – тельце Руффини (тепло).
Однако абсолютно «свободных» нервных окончаний среди тканевых элементов не существует, поскольку по данным электронной микроскопии все окончания в соединительной ткани окружены тонкими оболочками из шванновских клеток, и поэтому их также следует рассматривать как несвободные.
Свободные нервные окончания определяются в многослойных эпителиях и представляют собой тонкие ветвления дендритов без глиальной оболочки, лежащие между эпителиальными клетками. Такие окончания называют ноцицептивными, отвечают они за рецепцию боли. Также свободные окончания воспринимают тепло и холод.
Несвободные нервные окончания представлены древовидными тонкими нервными волокнами, окруженными узкими ободками цитоплазмы шванновских клеток. В коже такие нервные окончания тоже воспринимают боль.
Инкапсулированные нервные окончания имеют сложно организо-
ванные многослойные оболочки, состоящие из глиальных и соединительнотканных клеток.
Инкапсулированные рецепторы соединительной ткани всегда образованы осевым цилиндром и глиальными клетками. Обязательно
164
рецепторы такого типа покрыты снаружи соединительнотканной капсулой.
Тельца Фатера-Пачини – наиболее распространенный тип инкапсулированных рецепторов, представляющих собой пластинчатые тельца размером от 0,5 до 2 мм. Посредине рецептора расположена колба, состоящая из леммоцитов (рис. 62). При подходе к тельцу миелинизированное нервное волокно теряет миелиновую оболочку и погружается во внутреннюю часть тельца – луковицу. Наружная поверхность таких рецепторов покрыта капсулой из фибробластов и спиральных волокон. Воспринимаемыми сигналами являются тактильные и вибрационные. Находятся тельца Фатера-Пачини в глубоких слоях кожи, многочисленны в коже пальцев.
Тельца Мейснера (осязательные тельца) имеют размер 50– 150x60 мкм. Находятся в сосочковом слое
мененных нейролеммоцитов, окружены К инкапсулированным нервным цепторы мышц и сухожилий: нервно-
сухожильные веретена.
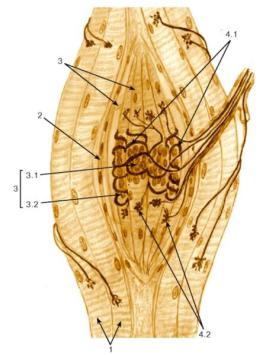
Нервно-мышечные веретена являются скелетных мышцах, которые функционируют жение. Веретено состоит из нескольких локон, заключенных в растяжимую интрафузальных волокон (рис. 63).
жащие за пределами капсулы, называются ла имеет слоистое строение. В ней различают слои. Между капсулой и интрафузальными полненное жидкостью пространство.
Интрафузальные волокна бывают двух
сумкой и волокна с ядерной цепочкой. ВолоконРис. 63. Нервнос ядерной-мышечноесумкверетеой в-
но (по В.Л. Быкову).
веретене у человека 1–3. В центральной расширенной части они со-
1 – экстрафузальные мышечные
держат много ядер. Волокон с ядерной цепочкой в веретене может
волокна, 2 – соединительнотканная
насчитываться 3–7. К интрафузальным мышечнымкапсула, 3 – интрафузальныеволокнам подхомы- дят афферентные волокна двух типов – первичныешечные волокнаи вторичные: 3.1 – мышечные. Пер-
волокна с ядерным мешочком (сум-
вичные волокна диаметром 17 мкм образуют окончания в виде спи-
кой), 3.2 – мышечные волокна с
рали – кольцеспиральные окончания – как на волокнах с ядерной
ядерной цепочкой, 4 – окончания
сумкой, так и на волокнах с ядерной цепочкойнервных волокон. Вторичные: 4.1 – кольцеспиволокна-
диаметром 8 мкм иннервируют волокна ральныес ядернойнервныецепочкойволокна. По, 4обе.2 –- им сторонам от кольцеспирального окончанигроздьевидныея они образуютнервные волокнагроздье-
видные окончания.
165
Интрафузальные волокна имеют также эфферентную иннервацию. К ним подходят тонкие моторные волокна, оканчивающиеся аксомышечными синапсами на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального волокна, они усиливают растяжение центральной рецепторной его части, повышая реакцию рецептора.
Нервно-сухожильные веретена обнаруживаются в месте соеди-
нения мышцы с сухожилием. Они служат для сигнализации о степени растяжения мышцы, состоят из нескольких сухожильных коллагеновых волокон, к которым подходят миелиновые волокна, затем теряющие миелин и разветвляющиеся в сухожильных волокнах. При растяжении импульс от нервно-сухожильных веретен идет в спинной мозг к тормозным нейронам, активность которых предотвращает перерастяжение мышцы.
2. Эффекторные нервные окончания
Эффекторные нервные окончания пронизывают все органы и ткани и оказывают на них влияние, кроме прямой передачи возбуждения на разные клетки, в первую очередь мышечные, эти окончания оказывают трофическое действие, регулируя метаболизм клеток и тканей.
Выделяют два типа эффекторных нервных окончаний – двига-
тельные и секреторные.
Двигательные нервные окончания представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Двигательные окончания поперечнополосатых мышц называются нервно-мышечными окончаниями (рис. 64).
В терминальных ветвях нервного волокна присутствует множество митохондрий и пресинаптических пузырьков с медиатором ацетилхолином.
Структурно нервно-мышечное двигательное окончание представляет собой своеобразный синапс, образованный пресинаптической мембраной, разветвлениями нервного волокна, синаптической щелью и постсинаптической мембраной – сарколеммой. Вся область контакта окружена нейролеммоцитами.
166
В гладкой мышечной ткани двигательные нервные окончания представлены своеобразными расширениями – варикозностями нервного волокна. Они по форме напоминают четки, внутри расширений содержится адреналин или ацетилхолин. Вблизи утолщений не обнаруживаются изолирующие клетки и волокна располагаются свободно.
Нейрожелезистые секреторные нервные окончания сходны по строению с нервными окончаниями в гладкой
мышечной ткани, образованы расширениями нервных волокон, в основном холинергичны.
3.Синапсы
Синапс – это контакт специализированных участков мембран нейронов, окруженный глиальной оболочкой, в котором осуществляется передача нервного импульса от одной клетки к другой. Синапсами также называют лишенные глиальной оболочки участки нейронов, через которые происходит передача возбуждения на эффективные клетки, которые могут быть мышечными (нейромышечный синапс) или эпителиальными (нейроэпителиальный синапс). Термин «синапс» впервые применил Шеррингтон в 1897 году. Синапсы визуализировались при импрегнации солями серебра или при окраске метиловым синим по Догелю. Существование синапсов было подтверждено после появления метода электронной микроскопии в середине двадцатого столетия, хотя их наличие предполагал еще С. Рамон-и- Кахаль. На поверхности нервных клеток мозга человека имеется примерно 1018 синапсов при плотности 109/мм3.
167
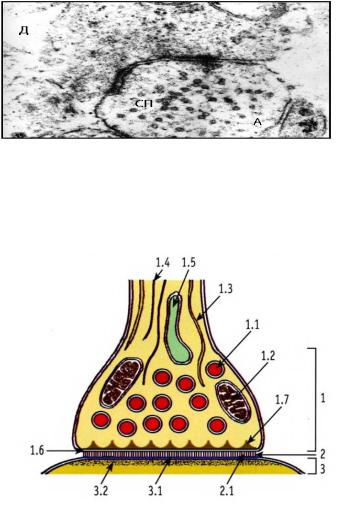
Классификация синапсов. В нервной системе описаны два вида синаптических контактов: 1) электрические синапсы и 2) химиче-
ские синапсы. Первые типичны для межнейронных связей в нервной системе низших позвоночных и для нейроглиальных клеток, формирующих своеобразные сплетения из астроцитов и олигодендроцитов. Вторые наиболее распространены в нервной системе млекопитающих, в том числе приматов и человека.
1.Электрический синапс
аналогичен по строению нексу- |
Рис. 65. Ультраструктура межнейрональ- |
||||||
су – щелевому контакту. Соеди- |
ного синапса коры большого мозга (препа- |
||||||
рат А.В. Солонского). A – аксон, Д – дендрит, |
|||||||
нения |
такого |
типа |
позволяют |
||||
СП – синаптические пузырьки. Ув. x70000 |
|||||||
двум клеткам обмениваться ин- |
|
|
|||||
формацией через поры, образо- |
|
|
|||||
ванные |
белком |
коннексином. |
|
|
|||
Размер пор совпадает с размера- |
|
|
|||||
ми ионов или небольших моле- |
|
|
|||||
кул. Отверстия, формируемые |
|
|
|||||
коннексином, не закрываются и |
|
|
|||||
не меняют размер, поэтому ток |
|
|
|||||
ионов или других веществ через |
|
|
|||||
них может происходить постоян- |
|
|
|||||
но в обоих направлениях. Отли- |
|
|
|||||
чительная особенность указанно- |
|
|
|||||
го типа контактов состоит в том, |
Рис. 66. Схема строения химическог |
||||||
что передаваемый сигнал дости- |
|||||||
синапса (по С.И. Юшканцевой, В.Л. |
|||||||
|
|
|
|
|
|||
гает клетки-мишени практически |
Быкову). |
|
|||||
мгновенно, в отличие от химиче- |
1 – пресинаптическая часть: 1.1 – си- |
||||||
ских синапсов, |
где присутствует |
наптические |
пузырьки с нейромедиа- |
||||
тором, 1.2 |
– митохондрии, 1.3 – |
||||||
"синаптичнская |
задержка", обу- |
||||||
нейротрубочки, 1.4 – нейрофиламенты, |
|||||||
словленная химическими |
преоб- |
||||||
1.5 – цистерна гладкой ЭПС, 1.6 – пре- |
|||||||
разованиями веществ при |
пере- |
синаптическая мембрана, 1.7 – преси- |
|||||
даче импульса от клетки к клетке. |
наптическое уплотнение; 2 – синапти- |
||||||
ческая щель: 2.1 – интрасинаптические |
|||||||
2. |
Химические синапсы осу- |
||||||
филаменты; 3 – постсинаптическая |
|||||||
ществляют передачу информации |
|||||||
часть: 3.1 – постсинаптическая мем- |
|||||||
с помощью системы мембран и |
брана, 3.2 – постсинаптическое |
||||||
химических веществ – нейроме- |
уплотнение |
|
|||||
диаторов (рис. 65). Поступающие |
|
|
|||||
168
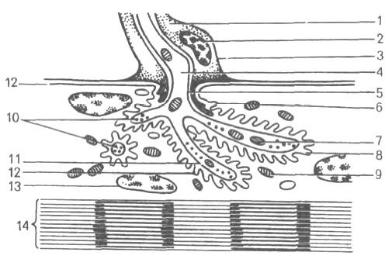
к синаптическим контактам электрические сигналы преобразуются в них в химические. Основными компонентами химического синапса являются: 1) пресинаптическая терминаль, 2) синаптическая щель, 3) постсинаптическая мембрана (рис. 66).
Пресинаптическая терминаль синапса имеет округлую или овальную форму, образована мембраной нейрона, от которого передается нервный импульс. К расширенной терминали подходят микротрубочки системы аксонального транспорта, по которым в область синаптического контакта перемещаются (белками-транслокаторами – кинезинами) пузырьки с нейромедиатором, а также микрофиламенты цитоскелета.
В терминали находятся единичные мембраны ЭПС, митохондрии для обеспечения энергией процесса выхода нейромедиатора из синаптических пузырьков и слияния их с пресинаптической мембраной. Этот процесс многостадиен и многокомпонентен, при этом он позволяет использовать мембранные структуры пузырьков многократно. Здесь же располагаются белки пресинаптического уплотнения. Приходящий к синапсу электрический импульс вызывает в пресинаптической терминали выброс ионов Ca2+, которые активируют присоединение пузырьков нейромедиатора к пресинаптической мембране. Молекулы нейромедиатора после слияния синаптических пузырьков и пресинаптической мембраны попадают в синаптическую щель.
Синаптическая щель – пространство между пресинаптической и постсинаптической мембранами шириной 10–50 нм. В пространстве синаптической щели находятся протеогликаны, связывающие мембраны между собой, – интрасинаптические филаменты. Диффундируя через синаптическую щель, молекулы нейромедиатора достигают постсинаптической мембраны. Однако не все молекулы участвуют в формировании постсинаптического сигнала – часть молекул нейромедиатора разрушается (моноаминоксидазой, ацетилхолинэстеразой и др.), часть подвергается обратному захвату в область пресинаптической мембраны (обратный захват серотонина), некоторое количество нейромедиатора выходит за пределы синаптической щели.
Постсинаптическая мембрана – часть синаптического контакта, принадлежащая клетке, воспринимающей информацию. На постсинаптической мембране находятся рецепторы, активация которых приводит к изменению активности контактирующего нейрона: либо его возбуждению, либо торможению, либо запуску в нем синтетиче-
169
ских процессов в зависимости от типа контактирующих клеток и типа нейромедиатора.
Синапсы подразделяют в зависимости от их расположения на нервных клетках на аксональные – аксо-дендритические, аксосоматические, аксо-шипиковые, аксо-аксональные; дендритические – дендро-соматические, дендро-дендритические, дендро-аксональные. В некоторых случаях устанавливаются контакты на той же самой клетке, от которой отходят отростки, такие контакты называют аутапсами.
Выделяют также контакты простые и сложные. Простые синапсы образованы одной пресинаптической и одной постсинаптической мембранами. Сложные синапсы включают несколько пресинаптических и синаптических областей, часто окруженных глиальной капсулой.
По степени изогнутости выделяют "положительно искривлен-
ные синапсы" – контактирующие мембраны выбухают в сторону постсинаптической мембраны, и "отрицательно искривленные синап-
сы" – мембраны изогнуты в сторону пресинаптической мембраны.
По степени выраженности пресинаптического и постсинаптического гранулярно-волокнистого парамембранного материала си-
напсы подразделяются на контакты с выраженным постсинаптическим уплотнением и широкой синаптической щелью (I, асимметричный, тип по Е. Грею, 1973) и синапсы со слабо выраженным постсинаптическим уплотнением и узкой синаптической щелью (II, симметричный, тип по Е. Грею). Также синапсы подразделяют на простые неперфорированные, простые перфорированные и множественные перфорированные.
По преобладающему нейромедиатору синапсы подразделяются на гистаминергические, адренергические, холинергические, серотонинергические, аминокислотные (таурин, глутамат, ГАМК, глицин), пептидергические, пуринергические.
По воздействию синапсов на клетку синапсы делятся на тормоз-
ные и возбуждающие.
Регенерация. Высокоспециализированные нейроны мозга взрослого человека теряют способность к митотическому делению и находятся в стадии G0 жизненного цикла клетки. Однако в последние годы появились доказательства неогенеза в некоторых отделах головного мозга, таких как гиппокамп, передний септум и обонятельная луковица. Выяснено, что стимулом к неогенезу в гиппокампе животных является систематическое применение антидепрессантов. В дру-
170