

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Физиология_центральной_нервной_системы
.pdfУчастие лабиринтов в тонических рефлексах продолговатого мозга было установлено Магнусом в следующих опытах. На шею кошки одевали гипсовую повязку, чтобы голова сохраняла по отношению к туловищу постоянное положение и проприорецепторы шеи не могли бы раздражаться. Животное вращали вокруг оси, проходящей через оба виска, и наблюдали. Если животное лежало спиной вниз, тонус разгибателей становился максимальным, а если спиной вверх, то тонус этих мышц уменьшался и конечности сгибались. Магнус пришёл к выводу, что эти рефлексы могли начаться только в рецепторах, которые чувствительны к изменениям земного притяжения, т.е. в рецепторах вестибулярного аппарата. При разрушении лабиринтов эти рефлексы не воспроизводились.
Адекватными раздражителями вестибулярного аппарата являются ускоренное или замедленное вращательное или прямолинейное движение. В начале движения головы эндолимфа в соответствующем полукружном канале отстаёт от движения головы, при этом отклоняются волоски рецептивных клеток в противоположном направлении, а в конце движения – по инерции продолжается движение, и волоски отклоняются от полукружных каналов. Происходит необходимое при этом или ином движении перераспределение тонуса и сохранение равновесия при движении.
Явления, связанные с двусторонним нарушением лабиринтов, обнаруживаются у человека лишь в течение первых месяцев после поражения. Возникает головокружение, тошнота, невозможность сохранять вертикальное положение тела. Восстановление нарушений, возникших после повреждения вестибулярного аппарата, происходит в результате зрения и проприорецепции. При одностороннем повреждении лабиринтов нарушения тяжелее и длительнее.
Значение проприорецепторов шейных мышц в позотонических рефлексах доказано в опытах с разрушенными лабиринтами. При различных поворотах головы происходит раздражение проприорецепторов мышц шеи и рефлекторное перераспределение тонуса в мышцах конечностей – разгибателях и сгибателях. Тонус разгибателей повышается с той стороны, куда повёрнута голова. При запрокидывании головы повышается тонус разгибателей передних конечностей.
61
Закономерности, найденные Магнусом у животных, необходимо учитывать в гимнастических упражнениях, при тренировке. В спортивной физиологии существуют принцип «ведущей роли головы». Движения тела облегчаются, если им предшествуют соответствующие движения головы.
Выпрямительные, или установочные, рефлексы осуществляются средним мозгом и отсутствуют у бульбарных животных. Выпрямительные рефлексы состоят из двух фаз: подъёма головы и рефлекторного выпрямления туловища. Животное после перерезки ствола мозга выше четверохолмия поднимает голову и встаёт на ноги. В этих рефлексах принимают участие лабиринты, проприорецепторы шейных мышц и рецепторы поверхности кожи.
Если животное положить на бок, то оно поднимает голову вследствие раздражения рецепторов вестибулярного аппарата. Если разрушить вестибулярный аппарат, то голова выпрямляется в том случае, если имеет место одностороннее раздражение кожных рецепторов. Однако, если на симметричную поверхность положить доску с грузом, то голова снова падает, что и доказывает значение рецепторов кожи в выпрямительных рефлексах. Сначала животное устанавливает голову теменем вверх, при этом раздражаются проприорецепторы шеи, и в ответ на это раздражение сокращаются мышцы, выпрямляющие туловище. Если одеть на шею гипсовую повязку, то туловище будет выпрямляться благодаря импульсам, поступающим с кожной поверхности. Нарушение этого рефлекса также доказывается в опытах с наложением на симметричную половину туловища доски с грузом.
Таким образом, для выпрямления как головы, так и туловища существуют два механизма: раздражение вестибулярного аппарата и кожной поверхности, раздражение проприорецепторов мышц шеи и рецепторов кожной поверхности.
В этих выпрямительных рефлексах активное участие принимает красное ядро среднего мозга.
Статокинетические рефлексы возможны при обязательном участии среднего мозга и возникают при угловом или линейном ускорении, как при положительном, так и при отрицательном. Наблюдается нистагм головы и глаз. Сначала голова и глаза
62
медленно вращаются в сторону, противоположную движению, затем быстро возвращаются в нормальное положение и т.д. При движении тела происходит перераспределение мышечного тонуса туловища и конечностей. Если собака поднимает одну ногу, то усиливается тонус мышц всех остальных конечностей, т.е. возникает устойчивое положение.
Во всех этих рефлексах принимает участие ретикулярная формация ствола мозга, которая является важнейшим регулятором мышечного тонуса, функционального состояния спинного и головного мозга. Роль ретикулярной формации в ЦНС сравнивают с ролью регулятора в телевизоре. Не давая изображения, он может менять громкость звука и освещённость.
Ретикулярная формация ствола мозга
Ретикулярная формация получила название в связи с тем, что в ней клетки располагаются на значительном расстоянии друг от друга и разделены большим количеством волокон, идущих во все направления. Нервные клетки неоднородны: их тела и отростки имеют разную длину и толщину. Дендриты нейронов ретикулярной формации тянутся на большом протяжении и дают сравнительно небольшое количество ветвей. Аксоны сильно ветвятся. Ретикулярная формация проходит через продолговатый мозг, мост, средний мозг и заканчивается в неспецифических ядрах таламуса.
Ретикулярная формация – это комплекс диффузно расположенных нейронов спинного мозга и ствола головного мозга, имеющих обширные связи между собой, с различными нервными центрами и корой головного мозга. Впервые было описана в 1880 г. О. Дейтерсом. Затем ее подробно описали Бехтеров и Рамон-и-Кахаль. Она напоминает сеть нервных образований, представлена рассеянными клетками в спинном мозге, продолговатом мозге, варолиевом мосту, среднем (покрышке), промежуточном мозге (субсталамическое ядро, гипоталамус). Ретикулярная формация имеет обширные двусторонние связи (сама получает и посылает импульсы). Получает она импульсы по коллатералям от восходящих путей. Эти импульсы поддерживают ретикулярную формацию в активном состоянии, ее тонус
63
(это неспецифическая афферентная система, потому что получает импульсы от разных раздражителей).
Основную роль выполняет ретикулярная формация ствола мозга. Ретикулярная формация занимает центральную часть на уровне продолговатого мозга, моста, среднего и промежуточного мозга.
Ретикулярная формация – это преимущественно неспецифическая часть ЦНС, т.е. основная масса ее нейронов и их проводников не являются центрами реализации определенных рефлексов. И тем не менее ретикулярная формация играет исключительно важную роль в осуществлении своих рефлекторных функций всеми ядрами ЦНС. Ее также называют «энергетической станцией ЦНС».
Ретикулярная формация мозга представлена сетью нейронов с многочисленными связями между собой и всеми структурами ЦНС. Она образует многочисленные афферентные пути: спиноретикулярный, церебеллоретикулярный, корково-подкорково- ретикулярный (от коры, базальных ганглиев, гипоталамуса), от структур каждого уровня ствола мозга (от среднего мозга, варолиева моста, продолговатого мозга), так и эфферентные нисходящие: ретикулоспинальные, ретикуло-корково-подкорковые, ретикуло-мозжечковые, к другим структурам ствола мозга. Таким образом, интенсивно обменивается чувствительной, так и двигательной импульсацией со всеми центрами ЦНС.
Ретикулярная формация оказывает генерализованное, тонизирующее, активирующее влияние на передние отделы головного мозга и кору больших полушарий (восходящая активирующая система ретикулярной формации), и нисходящее, контролирующее деятельность спинного мозга (нисходящая ретикулоспинальная система).
Ретикулярная формация регулирует ряд вегетативных функций, в реализации которых принимают стартовые нейроны ретикулярной формации, запускающие процесс возбуждения внутри нейронов, отвечающих за дыхательные и сосудодвигательные функции. В ретикулярной формации продолговатого мозга расположены два ядра, одно из них отвечает за вдох, другое – за выдох – дыхательный центр. Их деятельность контролируется пневмотаксическим центром ретикулярной формациимо-
64
ста. Повреждение дыхательного центра (центральный механизм дыхания) приводит к остановке дыхания.
Ретикулярная формация участвует в регуляции вегетативных функций эмоциональных реакций через гипоталамус, вегетативные ядра черепных нервов и лимбическую систему.
Сосудодвигательный центр расположен в ромбовидной ямке IV желудочка, входящего в состав ретикулярной формации. Если же в эксперименте перерезать ствол мозга сразу же за продолговатым, то у животного резко падает артериальное давление, развивается коллапс. Было выявлено две группы нейронов. Нейроны одной группы усиливают свою активность при повышении артериального давления, а нейроны другой – при его снижении.
Ретикулярная формация оказывает влияние на сенсорные системы мозга: на остроту слуха, зрения, обонятельных ощущений. Ретикулярная формация обладает высокой чувствительностью к таким веществам, как адреналин и углекислый газ.
В ретикулярной формации продолговатого, среднего мозга имеются нейроны, реагирующие на болевые раздражения от мышц и внутренних органов, при этом создается ощущение тупой боли.
Ретикулярная формация регулирует вегетативные и соматические реакции, условно-рефлекторную и психическую деятельность.
Установлено, ретикулярная формации имеет большое значение в регуляции возбудимости и тонуса всех отделов ЦНС, т.е. оказывает восходящее и нисходящее возбуждающее и тормозящее влияние. Нисходящее влияние ретикулярной формации на спинной мозг может быть как активирующее, так и тормозящее.
Активирующее влияние ретикулярной формации на спинной мозг поступает по ретикулоспинальным путям. Они подавляют активность клеток Реншоу. Устранение активирующего и тормозного действия ретикулярной формации при высокой перерезке спинного мозга является одной из главных причин спинального шока.
Ретикулярная формация продолговатого мозга изменяет тонус мотонейронов мышц-разгибателей. Частично перекрещенный латеральный ретикулоспинальный тракт идет от ретику-
65
лярного гигантоклеточного ядра продолговатого мозга в спинной мозг, волокна этого тракта оканчиваются на его вставочных нейронах. В результате возбуждаются α- и γ-мотонейроны мышц-сгибателей конечностей и реципрокно с помощью клеток Реншоу тормозят α- и γ-мотонейроны мышц-разгибателей. Волокна не перекрещенного медиального тракта идут от каудальных и оральных ретикулярных ядер моста и тоже оканчиваются на интеронейронах спинного мозга. Эти волокна возбуждают α- и γ-мотонейроны мышц-разгибателей, а через клетки Реншоу тормозят α- и γ-мотонейроны мышц-сгибателей.
Ретикулярная формация изменяет активность γ-мотонейро- нов спинного мозга, что является причиной тонуса мышц, перерезка задних корешков приводит к исчезновению тонуса. Осуществляется контроль за положением тела в пространстве. Поддержание тонуса мышц в покое является одной из важнейших функций ретикулярной формации. Ретикулярная формация является как бы «настройщиком», поддерживающим мышцы в постоянном тонусе.
Тормозящее влияние ретикулярной формации на спинной мозг. Впервые тормозящее влияние раздражения ретикулярной формации открыто И.М. Сеченовым в опытах на лягушках (раздражение зрительных бугров кристалликами поваренной соли). Впоследствии во многих исследованиях было показано, что электрическое раздражение различных участков ретикулярной формации продолговатого мозга может тормозить и усиливать некоторые спинномозговые рефлексы. Большое значение имеет сила раздражения ретикулярной формации.
Восходящее влияние ретикулярной формации на кору больших полушарий может быть как активирующим, так и тормозным.
Активирующее влияние ретикулярной формации среднего мозга оказывает активизирующее влияние на кору больших полушарий. Если перерезать ствол мозга между верхними и нижними холмиками четверохолмия, то нарушается поступление в кору больших полушарий возбуждения по сенсорным системам – зрительной и обонятельной. На ЭЭГ регистрируются медленноволновые регулярные ритмы, характерные для сна. У животного нарушен контакт с внешним миром, не реагирует на световые и обонятельные раздражители.
66
Г. Мэгуни и Дж. Моруцци (1949) показали, что стимуляция некоторых участков ретикулярной формации вызывает на ЭЭГ потенциалы, характерные для бодрствования, т.е. медленные ритмы сменяются высокочастотными ритмами.
Тормозное влияние ретикулярной формации на кору больших полушарий. При раздражении верхних отделов ретикулярной формации мозгового ствола животное впадает в состояние глубокого слоя (центры сна – Бремера, Чанга, Гессе). При этом на ЭЭГ возникает реакция синхронизации ЭЭГ-рит- мов. Таким образом, ретикулярная формация участвует в формировании цикла «сна и бодрствования».
Ретикулярная формация ствола мозга, в отличие от неспецифических ядер таламуса, оказывает длительное и генерализованное активирующее влияние на кору больших полушарий. Она поддерживает кору в активном состоянии. Кора тоже влияет на ретикулярную формацию, но это уже волевое сознательное влияние.
Степень воздействия ретикулярной формации на кору больших полушарий головного мозга снижается под влиянием некоторых фармакологических веществ. Этим определяется снотворное и наркотическое действие барбитуратов. Во время сна ретикулярная формация осуществляет контроль за сознанием и эмоциями. Этим обусловлена, например, реакция спящей матери, которая из всех доносящихся до нее звуков чутко выделяет плач своего ребенка и мгновенно просыпается.
Ретикулярная формация стимулируется не только нервными влияниями с различных рецептивных полей, но и гуморально. Она чувствительна к гормонам, продуктам метаболизма и медикаментам. Введение гормона адреналина в кровь возбуждает ретикулярную формацию среднего мозга, а его восходящие стимулирующие влияния повышают функции коры больших полушарий. Увеличение парциального давления углекислого газа возбуждает ретикулярную формацию дыхательного центра и стимулирует дыхание.
Как показал П.К. Анохин, аминазин блокирует ретикулярную формацию. Снотворные вещества, например, производные барбитуратовой кислоты (барбамил, тиопентал натрия, гексенал и др.) угнетают через кровь ретикулярную формацию, снимая ее
67
восходящее стимулирующее влияние на кору большого мозга, что способствует смене фаз бодрствования и сна.
Таким образом, ретикулярная формация участвует в регуляции деятельности вегетативной, соматической и психической деятельности организма.
Физиология мозжечка
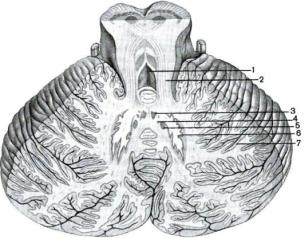
Мозжечок представляет собой образование, расположенное позади больших полушарий мозга над продолговатым мозгом и мостом мозга (рис. 18).
Он является отделом преимущественно соматического контроля, координируя сложные и автоматизированные движения, регулирует позу, движения, дополняя и исправляя деятельность остальных двигательных центров. В осуществлении этой функции важную роль играют обширные связи мозжечка с другими соматическими отделами ЦНС и рецепторным аппаратом. В мозжечок поступает информация от рецепторов мышц, суставов, сухожилий, кожи, вестибулярной, слуховой, зрительной систем.
Рис. 18. Строение мозжечка:
1 – IV желудочек; 2 – верхняя мозжечковая ножка; 3 – ядро шатра; 4 – шаровидное ядро; 5 – пробковидное ядро; 6 – зубчатое ядро; 7 – кора мозжечка
68
Выделяют три структуры мозжечка: древний мозжечок,
старый мозжечок, новый мозжечок.
Древний мозжечок – состоит из клочка, узелка и нижней части червя (язычка), имеет афферентный вход от вестибулярных ядер. Это объясняет значение мозжечка в регуляции равновесия.
Старый мозжечок – состоит из центральной дольки, вершины, пирамиды, язычка червя и четырехугольной дольки полушарий. Имеет афферентные входы от спинного мозга, от проприорецепторов мышц, сухожилий, надкостницы, оболочек суставов.
Новый мозжечок – состоит из полушарий и части червя (ската, листка и бугра); получает афферентные входы от разных областей коры, зрительных и слуховых сенсорных систем, участвует в анализе зрительных и слуховых сигналов и в организации соответствующих реакций.
Белое вещество мозжечка содержит три парных ядра (шатра, промежуточные и зубчатые), это главный эфферентный выход мозжечка на моторные центры головного мозга.
Кора мозжечка состоит из трех слоев и имеет специфическое гистологическое строение (рис. 19).
Верхний, или молекулярный, слой образован параллельными волокнами разветвлений дендритов и аксонов второго и третьего слоя. В нижней части расположены корзинчатые и звездчатые клетки, обеспечивающие взаимодействие клеток Пуркинье.
Второй (средний) слой коры состоит из выстроенных в один ряд клеток Пуркинье (грушевидные клетки). Они имеют самую мощную в ЦНС дендритную систему (дендритное поле одной клетки Пуркинье может содержать до 60 тыс. синапсов). Аксоны этих клеток являются единственным путем, с помощью которого кора мозжечка передает информацию в его ядра и ядра структуры большого мозга. Вся информация, приходящая в мозжечок, передается клетками Пуркинье. Они тормозят ядра мозжечка, а через них активность ретикулоруброспинальных нейронов, нейронов ядра Дейтерса, дающих начало вестибулоспинальному тракту.
Под клетками Пуркинье лежит третий слой – гранулярный, состоящий из клеток-зерен, число которых достигает 10 млрд. Их аксоны поднимаются вверх, образуя дорожки контактов с клетками Пуркинье, здесь же расположены клетки Гольджи.
69
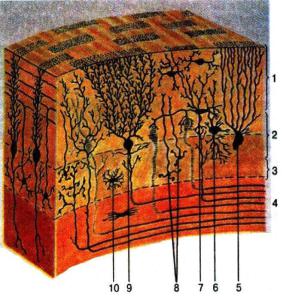
Рис. 19. Клеточные слои в коре мозжечка:
1 – молекулярный слой; 2 – слой грушевидных нейронов; 3 – зернистый слой; 4 – белое вещество; 5 – глиальная клетка; 6 – большая нервная клетка (клетка Гольджи); 7 – корзинчатая нервная клетка; 8 – малые нейроциты; 9 – ганглиозные нервные клетки (клетки Пуркинье); 10 – астроцит
Мозжечок связан с мозговым стволом тремя парами ножек: верхние соединяют его со средним мозгом, средние – с мостом, нижние – с продолговатым мозгом. Ножки мозжечка обеспечивают прямые и обратные связи с другими отделами мозга.
Верхние ножки – гладкая эфферентная система (зубчато- красноядерно-спинномозговой путь). Через верхние ножки информация идет в таламус, варолиев мост, красное ядро, ядра стола мозга, в ретикулярную формацию и кору больших полушарий, центральную извилину.
Через средние ножки к нему поступают двигательные сигналы от кортикоспинального (пирамидного) пути. Кора мозжечка обрабатывает эту информацию, затем через ядра мозжечка и его верхние ножки направляет двигательные импульсы к красным ядрам среднего мозга, который передает их вестибулярным ядрам и далее α-мотонейронам спинного мозга.
70