

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Физиология_центральной_нервной_системы
.pdfРуброспинальный (красноядерно-спиномозжечковый, пучки Монакова) путь состоит из аксонов нейронов красного ядра, расположенного в среднем мозге. Эти аксоны сразу после выхода из красного ядра переходят на симметричную сторону (перекрест) и разделяются на три пучка. Один идет в спинной мозг, другой – в мозжечок, третий – в ретикулярную формацию ствола мозга (продолговатый), заканчиваются на интернейронах соответствующего сегмента спинного мозга. Функции красных ядер – повышение тонуса сгибателей и понижение разгибателей. Значение в координации движений и тонуса.
Вестибулоспинальный путь начинается от нейронов ядра Дейтерса (латерального ядра), лежащего в продолговатом мозге. Это ядро регулирует активность мотонейронов спинного мозга, обеспечивает тонус мускулатуры, согласованность движений, равновесие и ориентацию в пространстве. Отвечает за формирование статических и статокинетических рефлексов, т.е. рефлексов поддержания позы при отсутствии или наличии ускорения. Примером рефлекса вестибулярных ядер может служить выставление рук в сторону падения тела. Этот рефлекс крайне устойчив, благодаря чему человек никогда не разбивается об опору даже при потере сознания.
Ретикулоспинальный (ретикулярно-спинномозговой) путь начинается на ретикулярных нейронах различных уровней моста и продолговатого мозга и заканчиваются на мотонейронах спинного мозга. Оказывает тормозное и облегчающее влияние на рефлексы спинного мозга. Отвечает за осуществление фазных двигательных реакций и поддержание позы тела.
Функция экстрапирамидного пути – осуществление точных автоматических движений (ходьба, письмо, езда на велосипеде, плавание, игра на музыкальных инструментах хорошо знакомой мелодии и т.п.).
Кортикоспинальные, или пирамидные, пути. Идут от круп-
ных пирамидных клеток, отдают большое количество коллатералей к ядрам промежуточного, среднего и продолговатого мозга. Различают двух видов:
1) прямые кортикоспинальные пути (пути передних столбов). В продолговатом мозге не перекрещиваются, идут в спинной мозг в составе вентрального пирамидного тракта и пере-
41
крещиваются в спинном мозге на уровне того сегмента, где они оканчиваются. У человека содержится 20%, у кошек – отсутствуют. Развитие идёт параллельно коре больших полушарий. Пирамидные пути на 80% состоят из волокон (группы С), проводящих импульсы из коры больших полушарий к мотонейронам передних рогов спинного мозга. Каждое полушарие иннервирует мускулатуру противоположной части тела;
2) кортикоспинальные пути боковых столбов. Перекрещиваются на уровне нижней трети продолговатого мозга. Оба пирамидных пути в верхних шейных сегментах содержат в два раза больше волокон, чем в грудных. Перерыв пирамидных путей приводит к нарушению произвольных движений. Поражение пирамидных путей ведёт к патологическим рефлексам.
Пирамидный тракт обеспечивает связь нейронов двигательной зоны коры больших полушарий с мотонейронами передних рогов спинного мозга и отвечает за произвольные, т.е. осознанные, движения. Другой особенностью пирамидной регуляции соматических движений является быстрота их реализации (низкая центральная задержка) при не очень высокой точности (так как участвует только один центр – это пирамидные нейроны Бетца прецентральной извилины коры головного мозга). Пример, удар рукой или ногой по мячу, наклоны корпуса для предотвращения столкновения.
Рефлекторная деятельность спинного мозга. В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются как соматические, так и вегетативные функции организма. Рефлекторную функцию спинного мозга обеспечивают афферентные и эфферентные нейроны, объединенные в рефлекторные дуги.
Нервные центры спинного мозга являются сегментарными рабочими центрами, обеспечивают сгибательные и разгибательные рефлексы. Нейроны спинного мозга непосредственно связаны с рецепторами и рабочими органами. Двигательные нейроны спинного мозга иннервируют все мышцы туловища, конечностей, шеи, а также дыхательные мышцы – диафрагму и межреберные мышцы. Надсегментарные рефлексы осуществляются только с помощью сегментов шейного отдела.
42
Рефлексы спинного мозга различают по рецепторам, при раздражении которых возникают рефлексы.
Проприорецепторные рефлексы возникают с мышечных,
сухожильных рецепторах, рецепторах надкостницы и оболочек суставов и участвуют в формировании ходьбы и регуляции мышечного тонуса.
Интерорецептивные рефлексы возникают с механо-, осмо-,
термо-, хеморецепторов и участвуют в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Кожные рефлексы возникают с экстерорецепторов и вызывают оборонительный защитный рефлекс.
Кроме того, спинальные рефлексы делят по рабочим органам:
1)рефлексы конечностей, которые могут быть фазными и тоническими;
2)брюшные рефлексы;
3)рефлексы в области таза.
Центры спинного мозга участвуют в регуляции большинства внутренних органов. Центр зрачкового рефлекса локализуется в сегментах (С8–Th2), регуляция деятельности сердца (Th1–Th2), слюноотделения – в сегментах (Th2–Th5), регуляция функций желудка – в сегментах (Th6–Th9), почек – в сегментах (Th5–L3). Из сегментов S2–S4 все органы малого таза (мочевой пузырь, часть толстой кишки ниже его левого изгиба, половые органы) получают парасимпатическую иннервацию. Сегментарно расположенные центры гладких мышц внутренних органов, центры пиломоторных рефлексов регулируют функции потовых желез и сосудов.
Во всех отделах спинного мозга находятся центры управления скелетной мускулатурой, иннервируют по сегментарному принципу. Мышцы шеи иннервируются С1–С4, сегменты С3–С5 иннервируют диафрагму, сегменты Th1–Th12 – наружные и внутренние межреберные мышцы, сегменты С5–Th2 – иннервируют руки, сегменты Th3–L1 – туловище, сегменты L2–S5 – иннервируют ноги.
Ретикулярная формация спинного мозга участвует в регуляции тонуса мышц, вегетативных функций, афферентной импульсации. Нейроны ретикулярной формации спинного мозга имеют большое количество отростков, обнаруживаются на
43
уровне шейных сегментов между передними и задними рогами, между боковыми и задними рогами в белом веществе, примыкающем к серому.
Соматические рефлексы спинного мозга. Соматические рефлексы бывают:
1)сгибательные (флексорные) в ответ на болевые разгибательные раздражители;
2)разгибательные (при надавливании на подушечки лапок);
3)рефлексы почесывания;
4)миостатические рефлексы, или рефлексы на растяжение мышцы, – при ударе по сухожилию происходит растяжение мышцы. α- и γ-мотонейроны поддерживают тонус тела и обеспечивают рефлексы сгибания и разгибания. У спинального человека эти рефлексы усилены.
1. Сгибательные рефлексы конечностей направлены на избежание различных повреждающих воздействий. Поэтому рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути.
Сгибательный рефлекс возникает при однократном раздражении болевых рецепторов кожи, мышц, внутренних органов. Вовлекаемые при этих раздражениях афферентные волокна имеют широкий спектр скоростей проведения – от миелиновых волокон типа А до безмиелиновых волокон типа С. Все афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, называются афферентами сгибательного рефлекса.
Рефлексы, возникающие с рецепторов кожи, являются полисинаптическими, они имеют защитное значение. Одновременно с возбуждением мотонейронов мышц-сгибателей происходит реципрокное торможение мотонейронов мышц-разгибателей.
В клинике исследуют такие сгибательные рефлексы, как локтевой, ахиллов (это проприорецептивные рефлексы) и подошвенный (кожный рефлекс).
2. Разгибательные рефлексы возникают только с проприорецепторов мышц-разгибателей, являются моносинаптическими.
3. Рефлексы почесывания – многократное повторное сгибание
иразгибание, т.е. происходят ритмические рефлексы конечностей.
44
Слабое раздражение кожи боковой поверхности тела у спинальной собаки вызывает почесывание этого участка задней конечностью, т.е. возникает чесательный рефлекс. Аналогом чесательного рефлекса у земноводных является потирательный рефлекс.
Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, которые устанавливают конечность в определенное положение к кожной поверхности.
Позные рефлексы, или рефлексы положения, представляют собой большую группу рефлексов, направленных на поддержание определенной позы. Примерами позного рефлекса являются сгибательные и разгибательные тонические рефлексы. Их можно вызвать при длительном растяжении мышц. Тонические рефлексы на растяжение мышц называют миотатическими.
Кроме того, спинной мозг выполняет следующие вегетативные рефлексы:
1)регуляция кровяного давления и сердца;
2)секреция и моторика желудочно-кишечного тракта;
3)регуляция функций мочеполовой системы;
4)потоотделение;
5)дефекация.
Спинальный шок. Рефлексы спинного мозга изучают на спинальной лягушке. При поперечной перерезке между спинным и продолговатым мозгом получают спинальную лягушку. Ниже место перерезки или травмы спинного мозга возникает явление спинального шока, выражающееся в резком падении возбудимости и угнетении деятельности рефлекторных функций всех центров спинного мозга, расположенных ниже места перерезки. В то же время деятельность центров, расположенных выше перерезки, сохраняется.
После перерезки исчезают не только скелетно-моторные рефлексы, но и вегетативные. Исчезают двигательные рефлексы, падает кровяное давление, отсутствуют сосудистые рефлексы, акты дефекации и мочеиспускание.
Причины спинального шока:
1. Выключение вышерасположенных отделов головного мозга, оказывающих на спинной мозг активирующее влияние, в
45
котором большая роль принадлежит ретикулярной формации ствола мозга.
2. По мнению некоторых ученых, перевозбуждение всех центров во время травмы вызывает их торможение.
Доказательства:
1)шоку подвергаются только центры спинного мозга, лежащие ниже места перерезки;
2)перерыв нисходящих путей спинного мозга приводит к подавлению рефлексов за счет выключения большого количества возбуждающих влияний на α- и γ-мотонейроны и другие спинальные нейроны, а также за счет растормаживания тормозных связей;
3)возникающая через определенное время после исчезновения явлений шока компенсация спинальных функций может привести к гиперрефлексии, которая является следствием выпадения тормозящих влияний ретикулярной формации ствола мозга. У человека с травмой спинного мозга раздражение стопы в период гиперрефлексии вызывает отдергивание обеих ног, потоотделение, мочеиспускание и дефекацию;
4)атравматическая перерезка спинного мозга, новокаиновая блокада, холодовая перерезка также вызывают спинальный шок;
5)повторная перерезка спинного мозга ниже первой повторно шок не вызывает.
Глубина и продолжительность шока различна у животных, стоящих на различных ступенях эволюционной лестницы. У лягушки шок продолжается 3–5 мин, у собаки – 7–10 дней, у обезьяны – больше 1 мес., у антропоидных обезьян и человека – несколько месяцев. Шок у человека нередко наблюдается как последствие бытовых или военных травм. Когда шок проходит, рефлексы восстанавливаются.
46
ГЛАВА 6 Общая характеристика головного мозга
Головной мозг является главным отделом ЦНС, выполняющим высшую регуляцию двигательных, чувствительных, висцеральных, эндокринных функций и психофизиологических процессов.
В среднем масса головного мозга составляет 1375 г у мужчин, 1245 г – у женщин, у новорожденных – 330–340 г. К 20 годам достигает окончательной величины.
Работами по изучению вариабельности строения мозга показано, что его масса не свидетельствует о состоянии интеллекта человека. Так, масса мозга И.С. Тургенева была 2012 г, а другого выдающегося писателя А. Франса – только 1017 г.
Головной мозг, также как спинной, состоит из серого и белого вещества, однако в головном мозге они устроены более сложно. Для ствола мозга характерно отсутствие метамерности, свойственной спинному мозгу. Серое вещество представлено ядрами в толще белого вещества, а у мозжечка и больших полушарий расположено по периферии и образует кору. Серое вещество определяет функцию того или иного отдела мозга. Белое вещество – нервные волокна (отростки нервных клеток), образующие проводящие пути (восходящие и нисходящие).
Австрийский анатом и основатель френологии Ф.Й. Галль впервые мозг разделил на три главных этажа: нижний – продолговатый мозг – «орган жизненных процессов»; средний – подкорка – «орган склонностей и влечений»; верхний – кора полушарий – орган интеллектуальных качеств души». Он составил карту черепа, где кружочками и цифрами обозначил расположение так называемых органов, например, «орган кровожадности», «знаменитая математическая шишка», «орган осторожности», шишка № 3 – «орган остроумия». Галль похоронен в Париже без головы, которую завещал для пополнения своей коллекции. Френология дала толчок появлению науки антропологии.
Головной мозг подразделяется на три основных отдела: мозговой ствол, мозжечок, конечный мозг (полушария большого мозга).
47
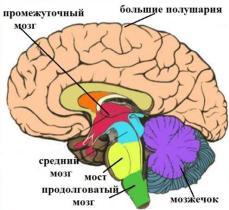
Ствол мозга
Ствол мозга продолжает спинной мозг вверх, в полость черепа, и состоит из продолговатого мозга, моста и среднего мозга (снизу вверх), промежуточного мозга (рис. 16).
Рис. 16. Основные отделы головного мозга
Функции ствола мозга
1.Осуществляет рефлекторные примитивные соматические рефлексы, направленные на поддержание позы тела в покое и при движении.
2.Обеспечивает различные вегетативные рефлексы.
3.Проводниковые функции. Через ствол проходят восходящие и нисходящие пути, связывающие кору больших полушарий со спинным мозгом.
4.Ассоциативные функции: связь различных отделов головного мозга между собой.
Другими словами, ствол мозга, как и любой другой отдел центральной нервной системы, выполняет две основные функции: рефлекторную и проводниковую.
48
Продолговатый мозг
Продолговатый мозг является частью головного мозга, соединяющей его со спинным мозгом. Его строение во многом сходно со строением спинного мозга, но в продолговатом мозге отсутствует сегментарное строение, серое вещество образует отдельные скопления нейронов – ядра. Центральный канал спинного мозга, заполненный цереброспинальной жидкостью, на уровне продолговатого мозга переходит в полость четвертого желудочка головного мозга. На вентральной поверхности четвертого желудочка располагается ромбовидная ямка, которая является ориентиром месторасположения ряда жизненно важных центров.
Выполняет две функции – рефлекторную и проводниковую.
Проводниковая функция продолговатого мозга. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спиноталамический, кортикоспинальный, руброспинальный, связывающие спинной мозг с вышележащими отделами ЦНС, и проводящие пути самого продолговатого мозга.
Впродолговатом мозге начинаются нисходящие пути – кортикоспинальные (пирамидные), которые соединяют крупные пирамидные клетки двигательной зоны с мотонейронами спинного мозга. Руброспинальный путь соединяет красное ядро с мотонейронами спинного мозга.
Собственно проводящие пути – вестибулоспинальный, оливоспинальный, которые связывают ядра вестибулярного нерва и оливу с мотонейронами спинного мозга. Служат эфферентными путями к мотонейронам спинного мозга при осуществлении вестибулярных рефлексов.
Внем заканчиваются пути из коры больших полушарий – корковоретикулярный, а также восходящие пути проприорецептивной чувствительности – тонкий пучок Голля и клиновидный пучок Бурдаха. Восходящие пути проприорецептивной чувствительности из спинного мозга заканчиваются около клеток ядер Голля и Бурдаха. Отростки клеток, находящиеся в этих ядрах, направляются в зрительные бугры, а затем в кору больших полушарий. Через продолговатый мозг проходят спиноталамические пути (латеральный и вентральный). Они проводят импуль-
49
сы болевой, температурной и тактильной чувствительности от клеток задних рогов спинного мозга к ядрам зрительных бугров. Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных пучками Голля и Бурдаха, а также находится ретикулярная формация.
Имеются также проводящие пути, соединяющие продолговатый мозг с мозжечком. К важнейшим ядрам продолговатого мозга относятся ядра Бехтерова и Дейтерса и нижняя олива. Ядра Бехтерова и Дейтерса связывают продолговатый мозг с мозжечком и красным ядром.
Центры продолговатого мозга. Собственно рефлекторная функция продолговатого мозга осуществляется расположенными в них нервными центрами. Здесь располагаются жизненно важные центры: дыхательный, сосудодвигательный, центр сердечной деятельности, пищеварительный, рвотный, кашлевой, чихательный. Поэтому не только удаление, но и повреждение продолговатого мозга заканчивается смертью.
Впродолговатом мозге выделяют центры защитных рефлексов: рвоты, кашля, чихания, слезоотделения, мигания и закрывания век. Здесь находится главный потоотделительный центр.
Впродолговатом мозге находится центр голосообразования, подчиненный центру речи в коре полушарий. При участии продолговатого мозга возможны лишь визг и нечленораздельные крики.
Большинство вышеперечисленных рефлексов продолговатого мозга реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга. От него идут команды к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного и подъязычного нервов,
врезультате чего реализуется тот или иной рефлекс. Вегетативные рефлексы реализуются здесь через располо-
женное в продолговатом мозге ядро блуждающего нерва, которое получает информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и затем ими же управляет. Ядро блуждающего нерва у
50