

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Физиология_центральной_нервной_системы
.pdfот начала раздражения до начала ответной реакции. Затем на область зрительных бугров накладывался кристалл поваренной соли, и время рефлекса увеличивалось, т.е. в таламической области мозга у лягушки существуют нервные центры, оказывающие тормозящее влияние на спинномозговые рефлексы.
Прекращение воздействия соли на зрительные бугры приводило к восстановлению исходного времени рефлекторной реакции.
Этот рефлекс осуществляется спинальными центрами, и его время служит показателем возбудимости нервных центров.
Кристаллик соли, раздражая зрительные бугры, вызывает возбуждение, которое распространяется к спинальным центрам и тормозит их деятельность. И.М. Сеченов пришел к заключению, что торможение является следствием взаимодействия двух или более возбуждений на нейронах ЦНС. В этом случае одно возбуждение становится тормозимым, другое – тормозящим.
Таким образом, И.М. Сеченов доказал, что наряду с торможением в периферической нервной системе существует и центральное торможение, характерное для ЦНС.
Торможение в ЦНС выполняет две основные функции. Во-первых, оно координирует функции, т.е. направляет возбуждение по определенным путям к определенным нервным центрам, при этом выключая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата. Если бы не существовало торможения, то каждый раздражитель, действующий на организм, вызывал бы реализацию своего рефлекса. В каждый конкретный момент времени на организм действует огромное количество раздражителей. В каждую секунду только в спинной мозг поступает такое количество чувствительной информации, которое соответствует всему объему информации, хранящейся в Российской государственной библиотеке. Если бы каждый из этих импульсов запускал рефлекс, то невозможно было бы осуществлять целенаправленную деятельность. Во-вторых, торможение выполняет охранительную, или защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей.
31
Торможение реализуется особыми тормозными нейронами ЦНС. Примерами таких нейронов могут служить клетки Реншоу спинного мозга, моховидные клетки мозжечка, тормозные нейроны ретикулярной формации.
Постсинаптическое торможение. В ЦНС наряду с возбуж-
дающими нейронами существуют тормозные нейроны. Аксоны этих нейронов образуют на телах и дендритах возбуждающих клеток нервные окончания, в которых вырабатывается тормозной медиатор глицин или ГАМК. Соответственно тормозные медиаторы взаимодействуют соответственными рецепторами. Например, глицин действует на глицинергические рецепторы, а ГАМК действует на ГАМКергические рецепторы. Нервные импульсы, возникающие при возбуждении тормозящих нейронов, не отличаются от нервных импульсов возбуждающих нейронов. Но в нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону импульса выделяется медиатор, который не деполяризует, а вызывает гиперполяризацию постсинаптической мембраны. Эта гиперполяризация регистрируется в форме электроположительной волны, обозначаемой как тормозной постсинаптический потенциал (ТПСП).
ТПСП ослабляет возбуждающий потенциал и препятствует достижению критического уровня деполяризации мембраны, необходимого для распространения потенциал действия.
Торможение называется постсинаптическим потому, что оно обусловлено гиперполяризацией постсинаптической мембраны.
Постсинаптическое торможение бывает:
1)параллельное;
2)латеральное;
3)возвратное;
4)прямое.
Пресинаптическое торможение возникает в аксо-аксо-
нальных синапсах. Локализуется именно в тончайших разветвлениях аксонов перед их переходом в нервное окончание. На этих терминалях располагаются окончания других нервных клеток, образующих здесь особые тормозные синапсы (рис. 14).
Медиаторы, выделяющиеся в этих тормозных синапсах, деполяризуют мембрану терминалей и приводят их в состояние,
32

подобно катодической депрессии Вериго. Это обусловливает частичную или полную блокаду проведения нервных импульсов к нервным окончаниям.
Пессимальное торможение.
Торможение в ЦНС может осуществляться и без участия особых тормозящих структур. В этом случае торможение развивается в возбуждающих синапсах в результате стойкой деполяризации постсинаптической мембраны под влиянием слишком частого поступле-
ния к ней нервных импульсов. Стойкая деполяризация постсинаптической мембраны нарушает распространение возбуждения, и рефлекс тормозится. К пессимальному торможению особенно склонны вставочные нейроны спинного мозга и клетки ретикулярной формации. Пессимальное торможение предохраняет нервную клетку от перевозбуждения.
33
ГЛАВА 5 Частная физиология ЦНС. Спинной мозг
Спинной мозг расположен в полости канала позвоночного столба. У плода позвоночный столб заполнен полностью, но к рождению он отстает только до нижнего края III поясничного позвонка, а у взрослого доходит лишь до II. Наиболее интенсивно спинной мозг растет до трех лет.
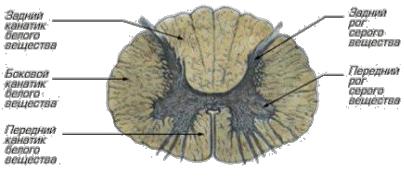
Спинной мозг состоит из серого и белого вещества (рис. 15). Серое вещество расположено внутри вокруг центрального канала. На поперечном разрезе напоминает летящую бабочку или букву Н. Серое вещество представлено преимущественно телами нервных клеток. Части серого вещества на поперечном разрезе называются рогами.
Рис. 15. Горизонтальный разрез грудного отдела спинного мозга
Передние рога по функции двигательные, в них находятся двигательные ядра, состоящие из соматических – двигательных клеток, волокна которых покидают спинной мозг в составе передних корешков.
Задние рога чувствительные, в них – чувствительные ядра, они состоят из вставочных клеток. Чувствительные клетки находятся в чувствительных ганглиях, их в ЦНС нет. В частности, в спинномозговых ганглиях, расположенных рядом со спинным мозгом в межпозвоночных отверстиях.
34
Боковые рога – есть только в нижних шейных, во всех грудных, в двух верхних поясничных сегментах – в них залегают вегетативные ядра симпатического отдела нервной системы.
Спинной мозг имеет сегментарное строение. Всего сегментов – 31–33, они соответствуют отделам позвоночного столба:
8 шейных (С1-С8), 12 грудных (Th1-Th12), 5 поясничных (L1-L5), 5 крестцовых (S1-S5), 1–3 копчиковых (Со1-Со3). Сегмент спин-
ного мозга – это поперечный срез спинного мозга с отходящими от него двумя парами корешков, которые, соединяясь, образуют периферические спинномозговые нервы. По закону Белла – Мажанди, задние корешки состоят из афферентных волокон, передние – из эфферентных. Перерезка задних корешков приводит не только к потере чувствительности, но и к нарушениям органов движения, которые становятся плохо координированными – атактическими. Спинномозговая атаксия возникает и у человека при поражениях восходящих путей спинного мозга.
От боковых борозд спинного мозга последовательно на всем его протяжении отходят корешки – передние и задние. Передний корешок спинномозгового нерва – двигательный, образован аксонами клеток двигательных ядер спинного мозга. Задний корешок спинномозгового нерва – чувствительный, образован аксонами чувствительных клеток спинномозгового узла.
Нейроны спинного мозга. Спинной мозг содержит около 13 млн нейронов, из них 3% – мотонейроны, 97% – вставочные, в том числе относящиеся и к ВНС.
Афферентные нейроны соматической нервной системы локализуются в спинальных ганглиях и ганглиях черепно-мозго- вых нервов. Их отростки в спинном мозге образуют синапсы либо на мотонейронах, либо на вставочных нейронах. По афферентным нервам в спинной мозг поступает возбуждение от экстеро- и проприорецепторов шеи, туловища, конечностей, а также от висцерорецепторов (кроме тех, что имеют связь с ЦНС через вагус).
Эфферентные нейроны спинного мозга относятся к соматической нервной системе, т.е. той, которая управляет деятельностью скелетных мышц. Это мотонейроны, или эфферентные нейроны.
35
Эфферентные нейроны спинного мозга представлены α- и γ-мотонейронами, расположенными в передних рогах спинного мозга, их аксоны образуют передние спинномозговые корешки, и вегетативными нейронами, находящимися в боковых рогах.
α-мотонейроны представляют собой крупные клетки с длинными дендритами. На соме и дендритах размещается до 10–20 тыс. синапсов. Аксоны характеризуются высокой скоростью проведения – 70–120 м/с. Они иннервируют экстрафузальные мышечные волокна (волокна скелетной мышцы), обеспечивая мышечные сокращение. Экстрафузальные мышечные волокна имеют диаметр 50–100 мкм, по длине варьируются от нескольких миллиметров до многих сантиметров. В связи с длительной следовой деполяризацией, сопровождающей потенциал действия, α-мотонейроны имеют низкую лабильность.
α-мотонейроны бывают: α1 и α2:
–α1-мотонейроны – иннервируют белые мышечные волокна, лабильность около 40 имп/с, их аксоны проводят возбуждение быстро;
–α2 -мотонейроны иннервируют красные мышечные волокна, аксоны проводят возбуждение медленно, лабильность этих волокон – 10–15 имп/с.
Аксон каждого мотонейрона делится на терминальные веточки, которые оканчиваются мионевральными синапсами на большом числе мышечных волокон, входящих в одну двигательную единицу. Совокупность мотонейронов, иннервирующих какую-либо мышцу, целиком образует мотонейронный пул.
γ-мотонейроны представляют собой значительно более мелкие клетки, иннервируют интрафузальные мышечные волокна, т.е. волокна мышечных веретен (рецепторы растяжения). Интрафузальные мышечные волокна имеют диаметр 15–30 мкм, длину 4–7 мм.
Несколько интрафузальных волокон сгруппированы и включены в соединительнотканную капсулу, названную мышечным веретеном, являющимся специализированным рецептором, отвечающим на растяжение мышцы.
Мышечные веретена располагаются параллельно экстрафузальным мышечным волокнам и удлиняются при растяжении мышцы.
36
Главная функция γ-мотонейронов состоит в предотвращении во время сокращения экстрафузальных волокон расслабления мышечных веретен. Возбуждение γ-мотонейронов способствует развитию начавшегося движения и используется для выполнения тонких и мелких движений. Они рассеяны среди α-мотонейронов и участвуют в регуляции мышечного тонуса. Их активность регулируется нейронами вышележащих отделов ЦНС. Следовая гиперполяризация в γ-мотонейронах выражена незначительно и не ограничивает существенно частоту их импульсации. В связи с этим они обладают высокой лабильностью (200 имп/с), аксоны проводят возбуждение медленно – 15–40 м/с.
Аксоны эфферентных нервов иннервируют все скелетные мышцы, кроме мышц лица.
Вегетативные нейроны. В боковых рогах с первого грудного сегмента и до первых поясничных сегментов спинного мозга находятся симпатические нейроны (С8-L3). Они вставочные, частота их разряда – 3–5 имп/с, относятся к двигательным нейронам, управляют деятельностью внутренних органов, органов головы, сосудов.
В боковых рогах серого вещества крестцовых сегментов находятся парасимпатические нейроны (S2-S4). Они тоже вставочные. Аксоны этих клеток выходят из спинного мозга в составе передних корешков спинномозговых нервов. Также относятся к двигательным нейронам, регулируют деятельности нижнего отдела толстой кишки и мочеполовых органов.
Интернейроны, или промежуточные нейроны, расположе-
ны в задних рогах и получают информацию от чувствительных нейронов. Следовая гиперполяризация в большинстве разновидностей вставочных нейронов выражена незначительно. Поэтому они способны к генерации высокочастотной импульсной активности. Им присуща высокая лабильность – до 400 имп/с. Они могут быть как возбуждающими и тормозными (клетки Реншоу, клетки Уилсона). В рефлекторной дуге эти нейроны являются вставочными (центральными). Тело, дендриты, аксоны находятся в пределах спинного мозга. Они обеспечивают ассоциативные связи спинного мозга.
Ассоциации реализуют в трех следующих направлениях:
1. Вставочные нейроны связывают афферентное и эфферентное звенья рефлекторной дуги.
37
2.Вставочные нейроны связывают между собой различные рефлекторные дуги данного и соседних (трех) сегментов спинного мозга.
3.Осуществляют связь спинного мозга с ядрами ствола мозга, а через них – с корой большого мозга.
Функции спинного мозга. Спинной мозг выполняет две функции: рефлекторную и проводниковую. Как рефлекторный центр спинной мозг способен осуществлять сложные двигательные и вегетативные рефлексы. Афферентными (чувствительными) путями он связан с рецепторами, а эфферентными – со скелетной мускулатурой и всеми внутренними органами.
Длинные восходящие и нисходящие пути спинного мозга соединяют двусторонней связью периферию с головным мозгом. Афферентные импульсы по проводящим путям спинного мозга приходят в головной мозг с информацией об изменениях во внешней и внутренней среде организма. По нисходящим путям импульсы от головного мозга передаются к эффекторным нейронам спинного мозга и регулируют их деятельность.
Проводниковая функция спинного мозга. В состав белого вещества спинного мозга входят миелиновые нервные волокна, собранные в пучки и образующие проводящие пути спинного мозга. Проводниковая функция осуществляется с помощью нис-
ходящих и восходящих путей. Короткие ассоциативные волок-
на обеспечивают межсегментарные связи, т.е. устанавливают связь между сегментами и внутри сегментов. Благодаря этим связям спинной мозг участвует в регуляции движений конечностей и туловища, тонуса мышц, обеспечивая поддержание позы.
Длинные проекционные волокна делят на восходящие, идущие к различным отделам головного мозга, которые получают информацию от тела организма и внутренних органов, управляют скелетной мускулатурой туловища и конечностей, регулируют деятельность внутренних органов и нисходящие – от головного мозга к спинному.
Восходящие пути (идущие к коре головного мозга) состоят из аксонов клеток спинальных ганглиев и пучковых клеток серого вещества. Проходят в белом веществе задних канатиков, расположенных между задними рогами спинного мозга. Соединяют сегменты спинного мозга со структурами головного мозга.
38
Функция этих путей заключается в передаче информации в вышележащие структуры ЦНС об экстеро-, интеро-, проприоцептивных раздражениях.
Тонкий пучок Голля отвечает за проведение проприоцептивной, тактильной, висцеральной чувствительности от нижней части туловища и нижних конечностей. Клиновидный пучок Бурдаха проводит проприоцептивную, тактильную, висцеральную чувствительность от верхней части туловища и верхних конечностей.
Оба пучка начинаются от рецепторов глубокой чувствительности мышц (проприорецепторов), сухожилий, надкостницы, оболочек суставов. Первый нейрон находится в спинномозговых ганглиях, а в задних столбах спинного мозга образуют синапсы на мотонейронах. Идут, не прерываясь в продолговатый мозг, где находятся тонкое и клиновидные ядра, происходит перекрещивание и синаптическое переключение на второй нейрон. Далее идут в латеральные ядра таламуса, где вновь переключаются на третий нейрон. Аксоны нейронов таламических ядер поднимаются и заканчиваются в IV слое соматосенсорной коры больших полушарий. Волокна этих трактов отдают коллатерали в каждом сегменте спинного мозга (заднецентральная извилина), что способствует коррекции позы всего туловища.
При поражении отмечается потеря тактильной чувствительности и нарушается координация движений.
Пучки Флексига и Говерса – спиномозжечковые пути. Формируются из аксонов интернейронов спинного мозга. Пучок Говерса также начинается нейронами спинномозговых узлов, затем переключается на клетки ядра промежуточной зоны. Аксоны переходят на противоположную сторону и поднимаются до верхних отделов ствола, где вновь перекрещиваются и вступают в ипсилатеральную половину мозжечка по его верхним ножкам, т.е. начинается на левой стороне тела и заканчивается в левой доле мозжечка. Пучок Флексига начинается нейронами спинномозговых узлов, переключается на клетках ядра Кларка. Аксоны не переходят на противоположную сторону и, достигая продолговатого мозга, через нижнюю ножку мозжечка вступают в ипсилатеральную (той же стороны) половину мозжечка. Этот путь начинается на правой стороне тела и заканчивается на правой доле мозжечка.
39
Эти пути передают импульсацию от проприорецепторов мышц и сухожилий, от висцерорецепторов к мозжечку. При перерыве этих путей происходит расстройство сложных движений и мышечного тонуса.
Спиноталамический путь – основной путь кожной чувствительности, начинающийся от болевых, температурных, тактиль-
ных рецепторов. Латеральный спиноталамический путь прово-
дит болевую и температурную чувствительности. Болевые, температурные и тактильные сигналы от рецепторов кожи идут в спинальные ганглии, затем через задние корешки к заднему рогу спинного мозга (первое переключение). Аксоны чувствительных нейронов переходят на противоположную сторону в каждом сегменте спинного мозга и поднимаются по боковому канатику к таламусу (второе переключение), а затем в сенсорную область коры большого мозга. Передний спиноталамический путь пере-
дает в зрительный бугор (таламус) тактильную чувствительность, не способен к тонкой дифференцировке локализации и интенсивности раздражителей, так как воспроизводит раздражение от большого количества рецепторов. Прерывается и перекрещивается в том же сегменте.
Нисходящие пути связывают отделы головного мозга с моторными или вегетативными эфферентными путями. Разделяются на несколько самостоятельных трактов, занимающих определенное положение в латеральных и вентральных канатиках белого вещества. Они идут от головного мозга к спинному. Нисходящие пути спинного мозга берут начало от нейронов, ядра которых расположены в пределах продолговатого мозга и моста – это ретикулоспинальный и вестибулярный тракты, а также от среднего мозга – руброспинальный тракт. Особое значение в соматической регуляции имеет путь, начинающийся от коры головного мозга – кортикоспинальный (пирамидный).
Соматические пути ЦНС делят на пирамидный и экстрапирамидный. Оба они заканчиваются на α- и γ-мотонейронах спинного мозга.
Экстрапирамидная система начинается от базальных ганглиев – подкорковых образований соматической нервной системы, и идет, прерываясь, в красных и вестибулярных ядрах, формируя, соответственно, рубро- и вестибулоспинальные тракты.
40