

Вторичная структура белка. Альфа- спираль и бета – складчатый слой.
Вторичная структура белков обусловлена способностью групп пептидной связи к водородным взаимодействиям: C=O. HN. При этом пептид стремится принять конформацию с образованием максимального числа водородных связей. Однако возможность их образования ограничивается тем, что пептидная связь имеет характер частично двойной связи и вращение вокруг нее затруднено, поэтому пептидная цепь приобретает не произвольную, а строго определенную конформацию (рис. 7). Известны несколько способов укладки полипептидной цепи в пространстве:
1) альфа-спираль — образуется внутрицепочечными водородными связями между NH-группой одного остатка аминокислоты и CO-группой четвертого от нее остатка (рис. 7А);
2) бета-структура (складчатый лист) — формируется водородными связями между пептидными группами полипептидных цепей, расположенными параллельно или антипараллельно (рис. 7Б), или связями между участками одной полипептидной цепи, образуя складки (рис. 7 В);
3) беспорядочный клубок — это участки, не имеющие правильной, периодической пространственной организации. Но конформация этих участков также обусловлена аминокислотной последовательностью.
Содержание альфа-спиралей и бета-структур в разных белках различно: у фибриллярных белков — только спираль или только складчатый лист; а у глобулярных белков — отдельные фрагменты полипептидной цепи: организованы в виде спирали либо складчатого листа, либо беспорядочного клубка.
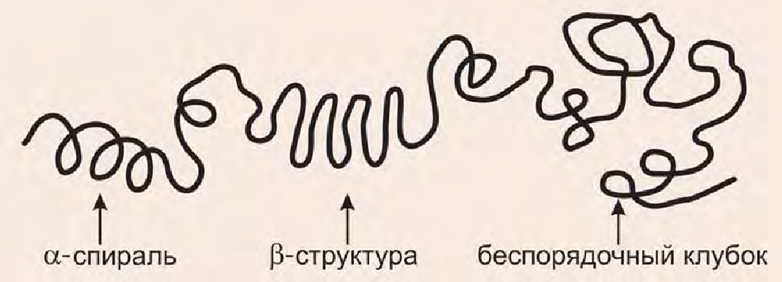
Рис. 7. Конформация полипептидных цепей:
А — α-спираль; Б — β-структура межцепочечная; В — структура в пределах одной пептидной цепи
Мобилизация гликогена при мышечной работе.
Контроль метаболизма гликогена осуществляется через регуляцию активности гликогенфосфорилазы (ключевой фермент распада гликогена) и гликогенсинтазы (ключевой фермент синтеза гликогена). Активация одного фермента сопровождается ингибированием другого.
В ответ на снижение концентрации глюкозы в крови синтезируется глюкагон, который стимулирует распад гликогена в печени. Запускается аденилатциклазный каскад реакций, приводящий к фосфорилированию и активации гликогенфосфорилазы и ингибированию гликогенсинтазы. Так же действует адреналин, связываясь с β2-рецепторами. Однако, если он связывается с α1-рецепторами, то запускается инозитолфосфатный механизм передачи сигнала. Результат действия обеих систем идентичен.
Гликогенолиз в скелетной мышце связан с участием Ca2+ в процессе мышечного сокращения. Кальций связывается с кальмодулином. Он активмрует гликогенфосфорилазу, и глюкозо-6-фосфат вступает в гликолиз, и обеспечивает образование АТФ, необходимого для мышечного сокращения.
При умеренной физической нагрузке и в состоянии покоя активатором гликогенфосфорилазы в мышцах является АМФ, образующийся при распаде АТФ.
В период пищеварения повышается концентрация глюкозы в крови, что является сигналом для синтеза и секреции инсулина. Под его влиянием ускоряется транспорт глюкозы в клетки инсулинзависимых мышечной и жировой тканей, активируется фосфопротеинфосфатаза, которая дефосфорилирует гликогенфосфорилазу, замедляя распад гликогена, и гликогенсинтазу, активируя её. Под действием инсулина индуцируется синтез глюкокиназы в печени, тем самым ускоряется фосфорилирование глюкозы. Глюкозо-6-фосфат аллостерически активирует гликогенсинтазу.
Так регуляция метаболизма гликогена в мышцах обеспечивает энергией интенсивную работу и энергозатраты в покое, а в печени поддерживает постоянство концентрации глюкозы в крови.