

4.2.3 Влияние взаимодействия междуRyR-каналами на стабильность осцилляций системы
Далее в ходе моделирования динамики Са2+-«часов» варьировался параметр взаимодействия между каналами в диапазоне от 0 до 2.5, и исследовалось влияние этого параметра на устойчивость осцилляций Са2+-«часов».
Результаты наблюдений авторитмических осцилляций изолированных внутренних Са2+-«часов» в виртуальном эксперименте представлены на рисунке 4.15.

Как показано на
рисунке, при увеличении параметра
![]() без учета взаимодействия нарушается
постоянство амплитуды зависимости
без учета взаимодействия нарушается
постоянство амплитуды зависимости![]() .
Эти нарушения проявляются в частотных
и амплитудных флуктуациях и внеочередных
высвобождениях, нарушающих гармонический
или релаксационный режим колебаний.
Под внеочередными высвобождениями
(обозначены на графиках красными
стрелками) подразумеваются высвобождения
при достаточно малой концентрации
.
Эти нарушения проявляются в частотных
и амплитудных флуктуациях и внеочередных
высвобождениях, нарушающих гармонический
или релаксационный режим колебаний.
Под внеочередными высвобождениями
(обозначены на графиках красными
стрелками) подразумеваются высвобождения
при достаточно малой концентрации![]() и, соответственно, при отрицательном
значении параметра эффективного давленияp,
как видно из временных зависимостей
p(t)
на вставках к графикам (рис.4.15).
и, соответственно, при отрицательном
значении параметра эффективного давленияp,
как видно из временных зависимостей
p(t)
на вставках к графикам (рис.4.15).
Данные внеочередные
высвобождения возникают вследствие
электронной активации RyR-каналов и
объясняются тем, что при достаточно
большой скорости заполнения СР
увеличивается частота высвобождений
и
![]() не успевает диффундировать в цитозоль
из диадного пространства. В связи с этим
вероятность электронных переходов не
успевает уменьшиться до нуля, инициируя
спонтанные высвобождения. Однако, как
видно из рисунка, наблюдается следующая
закономерность: при увеличении параметра
конформационного взаимодействия
соседних RyR-каналов уменьшается уровень
шумов и также количество внеочередных
всплесковCaSS
пример, при
не успевает диффундировать в цитозоль
из диадного пространства. В связи с этим
вероятность электронных переходов не
успевает уменьшиться до нуля, инициируя
спонтанные высвобождения. Однако, как
видно из рисунка, наблюдается следующая
закономерность: при увеличении параметра
конформационного взаимодействия
соседних RyR-каналов уменьшается уровень
шумов и также количество внеочередных
всплесковCaSS
пример, при
![]() =10
с-1
на первой секунде происходит восемь
внеочередных высвобождений при k=0,
в то время как при k=1
их число снизилось до двух. Следовательно,
усиление кооперативности привносит
стабильность осцилляций Са2+-«часов».
=10
с-1
на первой секунде происходит восемь
внеочередных высвобождений при k=0,
в то время как при k=1
их число снизилось до двух. Следовательно,
усиление кооперативности привносит
стабильность осцилляций Са2+-«часов».
Как было установлено экспериментально [126, 127], кооперативность RyR-каналов обеспечивает группа специфических белков FKBP 12.6, расположенных между каналами и стабилизирующих их динамику. При ослаблении действия данного белка с помощью различных препаратов было обнаружено нарушение самосогласованной динамики всего кластера [128], увеличение продолжительности локальных высвобождений Са2+ в диадное пространство (длительность спарков) и нарушения ритма сокращений [129, 130].
Изучение этих нарушений играет огромную роль в исследованиях возникновения аритмии. Внеочередные колебания внутренних Са2+-«часов», также как и флуктуации могут являться причиной нарушения автоволновой активности клеток водителя сердечного ритма в целом, что может стать аритмогенным фактором для всего миокарда.
Таким образом, на основе интеграции ЭК-модели RyR-каналов в модель кальциевой динамики можно сделать вывод, что введенная в рассмотрение кооперативная динамика RyR-каналов является стабилизирующим фактором, предотвращающим нежелательные сбои в активности клеток водителей сердечного ритма.
4.2.3 Эффект случайной остановки автоколебаний
На следующем этапе моделирования динамики Са2+-«часов» в настоящей работе исследовалось влияние скорости высвобождения Са2+ через одиночный RyR-канал на характер осцилляций системы.
Временные зависимости
относительного числа открытых каналов
и концентрации высвободившегося кальция
(![]() )
при различных значениях
)
при различных значениях![]() (от 1 до 100 с-1)
без учета и с учетом взаимодействия
между RyR-каналами показаны на рисунке
4.16.
(от 1 до 100 с-1)
без учета и с учетом взаимодействия
между RyR-каналами показаны на рисунке
4.16.

Эти зависимости
показывают, что без учета взаимодействия
при достаточно больших
![]() (>50 с-1)
наблюдаются устойчивые релаксационные
колебания Са2+-«часов»,
в то время как в интервале
(>50 с-1)
наблюдаются устойчивые релаксационные
колебания Са2+-«часов»,
в то время как в интервале
![]() от 10 до 50 с-1
проявляются как гармонические колебания,
так и спонтанные переходы к высокочастотным
стохастическим осцилляциям с малой
амплитудой вблизи определенного среднего
значения
от 10 до 50 с-1
проявляются как гармонические колебания,
так и спонтанные переходы к высокочастотным
стохастическим осцилляциям с малой
амплитудой вблизи определенного среднего
значения
![]() (~0.2 мкМ). Дальнейшее уменьшение параметра
(~0.2 мкМ). Дальнейшее уменьшение параметра![]() (
(![]() 5
с-1)
приводит к переходу осциллятора в
квазиравновесное состояние.
5
с-1)
приводит к переходу осциллятора в
квазиравновесное состояние.
При учете
взаимодействия между каналами наблюдается
стабилизация осцилляций концентрации
Са2+
в диадном пространстве при
![]() >15
с-1
(рис. 4.16б). Дальнейшее уменьшение параметра
>15
с-1
(рис. 4.16б). Дальнейшее уменьшение параметра
![]() приводит кспонтанным
переходам
осциллятора в стационарное
состояние,
то есть к постоянному току Са2+
из СР.
приводит кспонтанным
переходам
осциллятора в стационарное
состояние,
то есть к постоянному току Са2+
из СР.
Таким образом, в
численных экспериментах, проведенных
в данной работе, впервые наблюдался
принципиально новый эффект случайной
остановки Са2+-осциллятора.
Детальный анализ этого явления показал,
что он заключается в появлении в процессе
высвобождения/заполнения устойчивого
кластера открытых каналов
(2х2, 3х2 и проч.), через который и
осуществляется стационарное высвобождение
ионов Са2+
в диадное пространство. На рисунке 4.17
приведен пример данного перехода в
состояние стационарного высвобождения
на графиках зависимостей
![]() и
и![]() ,
и изображен вид решетки RyR-каналов при
переходе в данное состояние (светлыми
квадратами обозначены открытые, темными
– закрытые каналы).
,
и изображен вид решетки RyR-каналов при
переходе в данное состояние (светлыми
квадратами обозначены открытые, темными
– закрытые каналы).

Анализ этого рисунка позволяет сделать вывод, что формирование устойчивого кластера 3х2 RyR-каналов не является мгновенным, а происходит следующим образом: каналы, неокруженные открытыми соседями, при понижении CajSR в процессе высвобождения туннелируют в закрытое состояние. Вероятность туннелирования отрытых каналов с открытыми соседями достаточно мала, что приводит к появлению стабильного кластера в системе, стабильность которого обеспечивает конформационное взаимодействие между RyR-каналами.
Возвращаясь к
рисунку 4.16, можно заключить, что
необходимыми условиями для проявления
обнаруженного эффекта являются достаточно
сильное взаимодействие между соседними
каналами и высокий уровень критического
значения![]() по сравнению со средним значением
по сравнению со средним значением![]() .
В этом случае концентрация Са2+
в диадном пространстве не достигает
критического значения, в связи с этим
не происходят электронные переходы,
которые способны нарушить стационарность
системы.
.
В этом случае концентрация Са2+
в диадном пространстве не достигает
критического значения, в связи с этим
не происходят электронные переходы,
которые способны нарушить стационарность
системы.
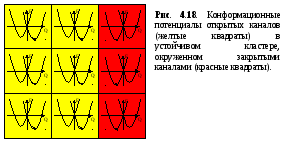
Для объяснения причин устойчивости открытого состояния каналов в кластере на рисунке 4.18 изображены конформационные потенциалы группы шести открытых каналов и соседних закрытых. Взаимодействие между открытыми каналами приводит к перенормировке конформационного потенциала в сторону стабильности открытого состояния. В связи с этим, канал, окруженный открытыми соседями, не может быть инактивирован даже при отрицательном значении параметра эффективного давления р, так как процесс туннелирования возможен только в случае глобального минимума обратного состояния.
В связи с тем, что случайные остановки Са2+-«часов» ранее не наблюдались, возникла необходимость более детального исследования данного эффекта, а также изучение влияния динамических параметров на размеры и устойчивость кластеров открытых каналов.