

Мускулатура головы филогенез мускулатуры головы
При описании скелетной мускулатуры туловища (в широком смысле* слова) и конечностей не было необходимости делить её на соматическую и висцеральную, так как почти вся она развивается из миотомов и их вентральных отростков, т. е. почти вся представляет соматическую мускулатуру.
Висцеральная мускулатура этих областей построена не из поперечнополосатой, а из гладкой мускульной ткани и распределена во внутренних органах, сосудистой системе, незначительным количеством—в основе кожного покрова.
Лишь очень небольшой пласт из поперечнополосатой мышечной ткани,, перемещающийся из области головы (бывшей жаберной области) на область шеи (трапецевидный мускул, передняя часть плече-головного мускула и некоторые другие), принадлежит по происхождению висцеральной мускулатуре,© чём & будет сказано ниже.
В области головы дело обстоит совсем иначе. Здесь и соматическая,-и большая часть висцеральной мускулатуры построена из поперечнополоса-ч той мускульной ткани, и лишь данные онто- и филогенеза, а также иннервация представляют более верный критерий для разграничения их между собой.
К соматической (париетальной) мускулатуре головы, происходящей от миотомов, относится мускулатура, управляющая движениями глазного яблока. Она иннервируется III, IV и VI парами черепномозговых нервов, (рис. 180—А, а, Ь, с, d, е, /).
Сюда же принадлежит мускулатура языка и подъязычной кости, иннервируемая XII черепномозговой парой. Она появилась в результате-диференцировки суббранхиальных (поджаберных), сегментально расположенных вентральных мускульных пластинок {А, 2). Ни та, ни другая в настоящем отделе не затрагивается: они описываются вместе с соответствующими органами (зрения, пищеварения).
Вся остальная мускулатура головы (и даже частью направляющаяся в отдел шеи) является висцеральной. Её можно разбить на две группы: а) одна из них ведает движением ротоглотки как начального этапа пищеварительного аппарата, а также гортани как участка дыхательных путей и иннервируется V, IX, X и XI парами черепномозговых нервов; б) другая представляет более поверхностный пласт и приводит в движение складки общего покрова головы: губы, щёки, ноздри, веки и ушные раковины. Эта группа стала, следовательно, кожной мускулатурой головы. Она иннервируется VII черепномозговой парой и по общему действию называется также мимической . мускулатурой.
Таким образом, с филогенетической точки зрения вся висцеральная мускулатура головы представляет результат сложной диференцировки метамер-ной жаберной мускулатуры (А, 3), иннервируемой V, VII, IX, X и XI парами черепномозговых нервов (указание на иннервацию, согласно сказанному на стр. 228 чрезвычайно важно для установления филогении и общей гомологии мускулов).
280
СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
V VH ix Х-ХП
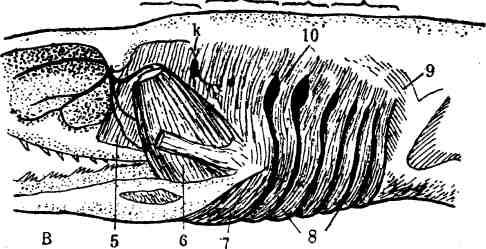
Уже у низших водных позвоночных метамерная жаберная мускулатура усложняется тем, что образует поверхностный сжиматель—constrictor super-ficialis,—который охватывает жаберный аппарат с вентральной и боковых сторон (В, 8). Впереди, в* пределах от челюстной дуги до местоположения брызгальца, этот мускул иннервируется двигательными ветвями V череп-номозговой пары (тройничный нерв) (В—V). От брызгальца до первой жаберной щели (пределы подъязычной дуги) мускул иннервируется ветвями VII черепномозговой пары (лицевой нерв) (В—VII). От первой до второй

Рис. 180. Л—схема распределения на голове соматической и висцеральной мускулатуры у низших позвоночных. В—распределение висцеральной мускулатуры головы
у акулы.
а, 6, с, d, е, /—мускулы, ведающие движением глазного яблока; д—орган слуха; h—продольная латеральная соединительнотканная перегородка между дорзальной и вентральной мускулатурой туловища; к—брызгальце; J, 2—боковая мускулатура (1—сё дорзальные мускулы, 2-—сё вентральные мускулы); 3—висцеральная мускулатура; 4—верхняя и нижняя челюсти; б—гёбноквадратный под-ниматель (m. levator paiatoquadrati); 6—приводитель вентральной челюсти (т. a Iductor mandibulae); 7—межчелюстной мускул (m. intermandibularis); 8—поверхностный сжиматель (m. constrictor super-iicialis); 9—примитивный трапецевидный мускул (m. trape7ir«s); 10—первая жаберная щель; V, \1U 7Х, X—XII—области иннервации (пятой» седьмой, девятой и десятой) черепномозгоЕЫми нервными
парами.
жаберной щели мускул получает нервы от IX пары—языко-глоточного нерва (В—IX). Наконец, от третьей жаберной щели до своего заднего края поверхностный сжиматель получает нервы от X (XI) черепномозговой пары (блуждающий нерв) и от не отделённого от него добавочного нерва (В—XI). Весь этот висцеральный мускульный пласт диференцируется на ряд отдельных мускульных единиц, из которых важно упомянуть лишь следующие:
1) Приводитель вентральной челюсти—m. adductor mandibulae (5, 6)— водных позвоночных, получающий двигательные ветви от V черепномозговой лары. У млекопитающих он даёт жевательные мускулы: большой жевательный мускул—m. masseter (рис. 188—10),—височный мускул—т. temporalis
ФИЛОГЕНЕЗ МУСКУЛАТУРЫ ГОЛОВЫ
281
(15)—и крыловой мускул—m. pterygoideus (рис. 184—а, 6). Кроме того, от этого мускула развивается напрягатель барабанной перепонки—m. tensor tympani.
Межчелюстной мускул—m. intermandibularis—водных позвоночных, получающий двигательные нервы от V черепномозговой пары; располагается он поперёк внизу между ветвями челюсти (рис. 180—В, 7). Он служит у наземных позвоночных материалом для развития подъязычно-челюстного мускула—m. mylohyoideus,—язычно-челюстного мускула—m. myloglos-sus,—а также переднего брюшка двубрюшного мускула—m. digastric us (рис. 184—в').
Приводитель подвеска—m. adductor hyomandibularis—и мускул жаберной крышки—т. opercularis,—обособляющийся у высших рыб; ин-нервируются они ветвями VII пары. У наземных позвоночных, ввиду значительного изменения прикрепления вентральной челюсти к черепу, вместо этих мускулов развивается опускатель вентральной челюсти—m. depressor mandibulae (иннервируется от VII пары). В частности, у млекопитающих опускатель вентральной челюсти усложняется и даёт яремно-подъязычный мускул—m. iugulohyoideus—и заднее брюшко двубрюшного мускула— т. digastricus,—которое у некоторых животных, например, у лошади, ответвляет ещё яремно-челюстной мускул—m. iugulomandibularis. К этой же мускульной группе, повидимому, принадлежит и стременной мускул—m. stapedius (см. орган слуха).
Мускульная масса жаберных дуг с исчезновением жабер у наземных животных (иннервируется IX и частью X парами) частью редуцируется, но в виде остатка диференцируется в мускулатуру подъязычной кости, глотки и гортани.
От того же поверхностного сжимателя, а именно от заднего участка области иннервации VII пары, происходит у рептилий довольно сильно развивающийся сжиматель шеи—m. sphincter colli,—охватывающий шею вентрально и с боков. У млекопитающих он распадается на два (даже три) пласта, которые служат материалом для диференцировки подкожной лицевой и шейной мускулатуры. В области головы из них развивается мимическая мускулатура, ведающая кожными складками (рис. 187).
Наконец, самый задний участок сжимателя представляет сравнительно незначительный трапецевидный мускул — m. trapezius, — который иннервируется задней частью X пары — добавочным нервом (рис. 180—5, 9).
Этот примитивный мускул у млекопитающих сильно разрастается и одновременно обособляется от X пары (блуждающего нерва), получая самостоятельную иннервацию от XI пары (добавочного нерва). Мускул далеко уходит назад и диференцируется на дефинитивный трапецевидный мускул— m. trapezius,—ключично-сосцевидный мускул—т. cleidomastoideus—и гру-динно-сосцевидный мускул—т. sternocephalicus; обычно последние два объединяются под одним названием—грудинно-ключично-сосцевидного мускула—m. sternocleidomastoideus.
С исчезновением у многих млекопитающих ключицы и изменением закрепления на черепе этот мускул получил название грудинно-головного мускула—m. sternocephalicus,—и плече-головного; мускула—т. brachio-cephalicus,—в состав которого вошла и ключичная часть дельтовидного мускула—pars clavicularis m. deltoidei,—и стал уже иннервироваться не черепномозговым, а спинномозговым нервом. Таким образом, этот мускул стал сложным (см. конечности).
Из группы мускулов, ведающих движениями складок кожного покрова головы, здесь рассматриваются только мускулы губ, подбородка, носа и щёк, а из группы мускулов ротоглотки и гортани—лишь жевательные.
282
СИСТЕМА ОРГАНОВ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
ОБЩАЯ ХАРАКТЕРИСТИКА ЛИЦЕВОЙ МУСКУЛАТУРЫ
Лицевая мускулатура представляет комплекс преимущественно пластинчатых мускулов, концентрирующихся вокруг имеющихся на голове отверстий. Их сложная диференцировка всё же подчиняется некоторой общей схеме. Вокруг каждого отверстия, сформированного кожными складками головы, мускульный пласт располагается в два слоя: один, обычно более глубокий, окружает отверстие кольцеобразно и составляет его запира-тель—с ф и н к т е р; другой, более поверхностный, распределяется ра-диально по отношению к отверстию и формирует его расширитель—д и л я-т а т о р.
А. Такое устройство чётко выражено вокруг отверстия рта. Неодинаковые образ жизни и способы захвата пищи обусловливают разнообразие устройства входа в ротовую полость, называемого ротовой щелью. Низшие наземные позвоночные—амфибии и рептилии—имеют обычно широкую1 ротовую щель, обрамлённую плотными складками кожного покрова. Такие примитивные губы лишены собственной мускулатуры, следовательно, сами по себе неподвижны. Они подчиняются движению челюстей.
Млекопитающие (за исключением однопроходных) обладают свисающими от области дёсен сильно выраженными сравнительно мягкими складками, которые с боковых сторон ротовой полости значительно надвигаются вперёд, образуя в области коренных зубов стенки, называемые щеками. Вследствие этого сама ротовая щель, по сравнению с таковой рептилий, укорачивается и располагается лишь в начальном (переднем) участке морды. Охватывающие её длинные складки представляют настоящие дефинитивные губы, способные к самостоятельным движениям, т. е. вне зависимости от работы челюстей. Таким образом, по происхождению щёки и губы представляют неразрывное целое.
Сфинктером ротовой щели служит заложенный в толщу губ круговой мускул рта—ш. orbicularis oris (рис. 157—4).
Дилятаторы расходятся радиально от ротовой щели к поверхностям соседних костей, причём среди них имеются более короткие пучки и более длинные отдельные мускульные ленты. Последние составляют группу обособленных мускулов и дают возможность осуществлять одностороннее, с большим размахом оттягивание одной из губ или даже её небольшого участка; к коротким пучкам принадлежат верхние и нижние резцовые мускулы— mm. incisivi,—идущие от резцовых частей челюстей в толщу губ. К длинным пучкам принадлежат: специальный подниматель верхней губы— m. levator labii superioris proprius (2),—носо-губной подниматель—m. levator nasolabialis (2),—клыковый мускул—m. caninus (5),—опускатель нижней губы—m. depressor labii inferioris (£),—кожный мускул губ—m. cuta-neus labiorum—и скуловой мускул—m. zygomaticus (4'). Из приведённой схемы видно, что все они стягиваются к губам, но некоторые из них подходят и к отверстиям носа.
Среди домашних животных подвижность губ далеко не одинакова. Лошади и мелкие жвачные обладают самыми подвияшыми губами (и способны легко выбирать наиболее приемлемое из питательного материала). Рогатый скот и свиньи имеют весьма мало подвижные губы, особенно свиньи. Естественно, что у последних эти мускулы крайне ослаблены в своём развитии и часть их (специальный подниматель верхней губы) приспособлена к движению'хоботка. Собаки также не обладают диференцированной подвижностью губ и лишь при гневе путём раскрывания ротовой щели оголяют резцовые зубы и клыки.
Мускулатура щёк в деталях распределения своих пучков довольно разнообразна даже среди домашних животных и представляет, повидимому,
ОБЩАЯ ХАРАКТЕРИСТИКА ЛИЦЕВОЙ МУСКУЛАТУРЫ
диференцировку общего сфинктера ротовой полости (Арсеньев). Более поверхностные пучки идут в толще щеки дугой от дорзальной челюсти к вентральной, а глубокие располагаются более продольно. Сюда относится щёчный мускул—m. buccinator,—имеющий поверхностный слой—m. buccalis— и глубокий слой—т. molaris. Оба слоя при жевании помогают^юдкладывать спадающую с коренных зубов пищу на их режущую (трущую) поверхность. Это достигается сжиманием, т. е. уменьшением, защёчного пространства, и по существу схоже с действием сфинктера ротовой щели. При режущей конструкции зубов, как, например, у собак, щёчная мускулатура построена примитивнее, при перетирающей конструкции у травоядных— сложнее.
Б. Носовые отверстия, называемые ноздрями, обладают довольно сложной дилятаторной мускульной группой, причём здесь, как вокруг ротового отверстия, имеются короткие дилятаторные пучки, которые по силе развития очень разнообразны, и длинные мускулы носа. Последние могут быть по работе одновременно и мускулами ротовой щели. Лищь у некоторых видов диких животных встречаются сфинктеры.
Наиболее хорошо развиты короткие дилятаторы у лошадей в виде поперечного мускула носа—m. transversus nasi (рис. 182—13)—и латерального мускула носа—m. lateralis nasi (9, 10, 11, 12). Последний имеет оральный, ■аморальный, дорзальный и вентральный пучки. Ноздри лошадей действительно способны весьма значительно расширяться при усиленных актах дыхания,'превращая сравнительно узкие щелеобразные отверстия, в широкие округлые входы в носовую полость.
У свиней и собак короткие дилятаторы или рудиментарны, или совсем не развиты. У рогатого скота они слабее, чем у лошадей.
Длинные дилятаторы ноздрей в главной массе представляют лишь пучки, отъединяющиеся от мускулов ротовой щели, как, например, пучки от носо-губного поднимателя—m. levator nasolabialis—и от клыкового мускула—m. caninus. У собак эти пучки подходят к подвижной части носа, расщепляются здесь на серию тонких веток, которые закрепляются вокруг ноздрей и при одновременном право- и левостороннем действии могут слегка расширять ноздри, а при одностороннем действии—поворачивать верхушку носа вправо или влево. Для этой же функции у собак приспособлен и специальный подниматель верхней губы — m. levator labii superioris proprius.
У лошадей и жвачных такие пучки служат для расширения ноздрей, а у свиней они используются для особого движения хоботка, к которому стягиваются пучки со всех сторон; они позволяют хоботку производить небольшие экскурсии в сагиттальной и фронтальной плоскостях. Для выполнения той же функции у свиней существует ещё специальный опускатель хоботка—m, depressor rostri.
В. В кожных складках, формирующих отверстие для органа зрения и называемых веками, распределение мускулатуры подчинено общей схеме, но в упрощённом виде, т. е. они имеют сфинктеры и дилятаторы (см. орган зрения).
Значительно многограннее диференцировка мускулатуры отверстия наружного уха, представляющего также складку общего покрова с вправленной хрящевой пластинкой—ушной раковиной. Её наличие и необходимость в особых оттенках подвижности обусловливают сложность распределения сфинктеральной, или частной, и дилятаторной, или общей, мускулатуры уха, но общие черты расстановки мускулов (сфинктер и дилятатор) вокруг данного рупорообразного отверстия всё же ясно заметны (см. орган слуха).
284 система органов произвольного движения
ОБЩАЯ ХАРАКТЕРИСТИКА ЖЕВАТЕЛЬНОЙ МУСКУЛАТУРЫ
Жевательная мускулатура не многочисленна, но мощна. Исходное неподвижное прикрепление—punctum fixum—всех этих мускулов находится в области мозгового отдела черепа, а конечное подвижное—punctunr mobile—на вентральной челюсти. Она одна только и является подвижной костью при жевании. Точкой опоры при её движении служит парный челю-стный сустав, который может увеличивать свой угол, т. е. разгибаться, и уменьшать его, т. е. сгибаться. Увеличение угла влечёт за собой размыкание челюстей, а уменьшение—смыкание их. Это и есть основное и наиболее обширное движение при жевании.
У свиней и особенно у собак такой вид движения является почти исключительным, так как зубы собак приспособлены для простого разрезывания,. а у свиней—для раздробления твёрдого питательного материала. Однако челюсть свиней может производить слабые боковые движения и выдвигание вперёд. У лошадей и жвачных, помимо дробящего действия, наблюдаются уже ясно выраженные боковые движения вентральной челюсти и более тщательное перетирание питательного материала, в соответствии с чем перестроены и коренные зубы. Кроме того, у них возможно лёгкое выдвигание вентральной челюсти вперёд.
Вентральная челюсть может быть рассматриваема в качестве рычага и первого и второго рода. Рычагом второго рода она бывает, когда плечом сопротивления служит участок челюсти от челюстного сустава вниз и вперёд до резцовых зубов, а плечом силы—челюстная ветвь и её венечный отросток, на который действует височный мускул—m. temporalis,—отходящий от височной ямки (рис. 157—<3), и веерообразно расправленные большой жеватель-. ный (7) и крыловой мускулы—m. masseter и т. pterygoideus. Первый отходит от скуловой дуги, а второй—от области крыловидного отростка клиновидной кости. Все три мускула при сокращении совместно смыкают челюсти,, и тем самым рычаги скорости приобретают огромную силу.
При отведении вентральной челюсти вбок у травоядных действует правый жевательный мускул совместно с левым крыловым, и челюсть отходит вправе, а при сокращении другой соимённой пары—влево. Этому у травоядных помогает анизогнатное устройство челюстей. При выдвигании вентральной челюсти вперёд сокращаются пучки большого жевательного и крылового мускулов, направленные от своего начала назад, к поверхностям .прикрепления на ветви вентральной челюсти.
При размыкании вентральная челюсть находится в условиях рычага первого рода с плечом силы от челюстного сустава до места закрепления двубрюшного и яремно-челюетного мускулов—m. digastricus et m. iugulo-mandibularis (#); плечом сопротивления служит участок челюсти от сустава до резцовых зубов. Этим мускулам у лошадей и жвачных помогает грудин-ко-головной мускул—m. sternocephalicus (10).
При обоих условиях рычаги используются в качестве рычагов скорости.
МУСКУЛАТУРА И ФАСЦИИ ГОЛОВЫ У ЛОШАДЕЙ ФАСЦИИ ГОЛОВЫ
С области шеи поверхностная и глубокая фасции переходят на отдел головы, где и формируют поверхностную и глубокую фасции головы.
1) Поверхностная фасция головы лежит непосредственно под кожей, одевает почти всю голову и содержит в себе кожную мускулатуру. В различных областях она получает самостоятельные названия. Околоушн о— жевательная фасци я—fascia parotideomasseterica—покрывает
МУСКУЛАТУРА ГОЛОВЫ
285
околоушную железу и жевательный мускул и закрепляется на скуловом гребне. Она содержит вентральный ушной мускул (см. орган слуха). П о-верхностная височная фасци я—fascia temporalis superficial—охватывает область височного мускула и заключает общие ушные мускулы этой области. Лобная фасци я—fascia frontalis—лежит в области лба и перехода в спинку носа. Щёчная фасци я—fascia buccalis—покрывает носовую область и щёчный мускул. Она включает в себя носо-губной подниматель, кожный мускул губ, скуловой мускул и опускатель нижнего века. Подчелюстная фасция —fascia submaxillaris—находится в подчелюстной области и содержит в себе часть подкожного мускула губ.
2) Глубокая фасция головы лежит глубже поверхностной и в области щеки тесно связана с ней. Она также в ряде мест имеет свои названия. Щёч-но-глоточная фасци я—fascia buccopharyngea—локализуется на медиальной поверхности брюшка большого жевательного мускула, откуда переходит вперёд, на область внутренней поверхности вентральной челюсти. Здесь она достигает слизистой оболочки щеки, прилегает к внутренней поверхности опускателя вентральной губы и щёчного мускула и отдаёт листок на наружную сторону, одевая эти мускулы и щёчные слюнные железы. Кверху она окружает клыковый мускул, подходит под специальный подниматель верхней губы и теряется в области носа, частично сливаясь с поверхностной фасцией. Назади эта фасция одевает мускулатуру глотки. Она фиксируется на крючке крыловидной кости и рядом на дорзальной челюсти, откуда в качестве крыловидн о-ч елюстной связки спускается к верхнему краю вентральной челюсти позади коренных зубов, а затем закрепляется на подъязычной кости. Глубокая височная фасция—fascia temporalis profunda—одевает непосредственно височный мускул, с перимизием которого тесно соединяется, а также находящийся снаружи орбиты глазной жир и закрепляется на скуловой кости и скуловом отростке лобной кости.