

Микробиология и основы биотехнологии
..pdfкулы АТФ превращается в тепловую, и энтропия, в соответствии со вторым законом термодинамики, возрастает. Однако в клетке концевая фосфатная группа в процессе гидролиза не просто отделяется, а переносится на особую молекулу, служащую акцептором (приемником). Значительная часть свободной энергии молекулы АТФ сохраняется благодаря фосфорилированию (присоединению одной фосфатной группы) молеку- лы-акцептора, которая за счет возросшей энергии вновь приобретает возможность участвовать в процессах, протекающих с потреблением энергии (процессы биосинтеза). В термодинамике клетки молекулу АТФ можно рассматривать как «заряженную» форму носителя энергии, а АДФ — как «разряженную» форму. Первичная «зарядка» АТФ происходит в процессе фотосинтеза, в котором энергия, отданная электронами, используется на образование АТФ из АДФ и фосфата, т. е. на «зарядку» аденозинфосфатной системы фотосинтезирующей клетки.
Вторичная «зарядка» производится в процессе дыхания, когда энергия, заключенная в питательных веществах субстрата, освобождается в результате окисления и расходуется на построение АТФ из АДФ и фосфата.
5.3. Фотосинтез
Роль АТФ в процессе фотосинтеза удалось выяснить в шестидесятых годах прошлого века. Это открытие позволило установить механизм связывания солнечной энергии фотосинтезирующей клеткой в процессе синтеза углеводов. Энергия солнечного света передается в виде фотонов, или квантов. Свет разной длины волны характеризуется различной энергией. В клетках растений и некоторых микроорганизмов, содержащих хлорофилл, солнечный свет с определенными длинами волн поглощается зеленым пигментом. Хлорофилл физиологически активен только в связанной с белком форме, выделенный препарат хлорофилла не способен использовать энергию света, которая быстро рассеивается.
Образование органического вещества в процессе фотосинтеза происходит в двух циклах.
Световой цикл фотосинтеза (цикл Д. Арнона)
Фотон света, попадая в молекулу хлорофилла, поглощается. Под влиянием поглощенного света молекула хлорофилла приходит в триплетное «возбужденное» состояние. Поглощенная энергия переводит
71
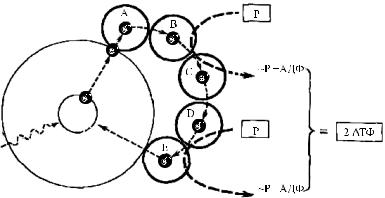
электрон в сложной молекуле хлорофилла с основного устойчивого энергетического уровня на более высокий энергетический уровень. Богатые энергией электроны не возвращаются в свое нормальное положение, а отрываются от молекулы хлорофилла и переносятся специфическими молекулами переносчиков электронов (А, В, С, Д, Е), которые передают электроны друг другу по замкнутой цепи. Проделывая этот путь вне молекулы хлорофилла, возбужденные электроны постепенно отдают свою энергию переносчикам и возвращаются на свои прежние места в молекуле хлорофилла, которая оказывается готовой к поглощению второго фотона. Энергия, отданная электронами, расходуется на образование двух молекул АТФ из двух молекул АДФ. Переносчики электронов, служащие посредниками в процессе фотосинтетического циклического фосфорилирования (процесс, при котором электрон в молекуле хлорофилла, перенесенный на более высокий энергетический уровень благодаря поглощению фотона света, обеспечивает энергию, необходимую для образования АТФ), еще не вполне установлены. Один из них, по-видимому, содержит рибофлавин (витамин В2) и витамин К. Другие предварительно отнесены к цитохромам — белкам, содержащим в своем составе атомы железа, окруженные порфириновыми группами.
Схема циклического фосфорилирования представлена на рис. 5.1 [18]. В процессе фотосинтеза, помимо связывания солнечной энергии, осуществляется и синтез углеводов. Для этого необходим внешний источник электронов и протонов водорода, что дает возможность синтези-
Рис. 5.1. Схема превращения солнечной энергии в энергию фосфатных связей АТФ в цикле Арнона
72
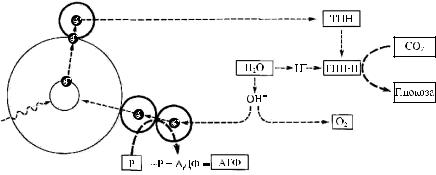
Рис. 5.2. Схема восстановления ТПН протонами Н+, полученными из воды
ровать углевод путем «восстановления» двуокиси углерода (т. е. добавления к ней электронов и протонов водорода).
Источником электронов служит хлорофилл, источником протонов — вода. Восстанавливающимся агентом служит трифосфопиридиннуклеотид (ТПН), который восстанавливается (ТПН·Н) под действием протонов и электронов. Отщепляющиеся при этом от молекулы воды ионы гидроксила (ОН–) отдают свои электроны цитохромам, которые передают их в ряде фотохимических реакций молекуле хлорофилла. В результате образуется газообразный кислород — характерный продукт фотосинтеза.
В настоящее время считают, что некоторые из «горячих» электронов возбужденной молекулы хлорофилла вместе с ионами водорода, происходящими из воды в процессе ее фотолиза, служат дополнительным источником электронов или атомов водорода, вызывающих восстановление ТПН. Схема восстановления ТПН протонами Н+, полученными из воды, представлена на рис. 5.2 [18].
Темновой цикл фотосинтеза (цикл М. Кальвина)
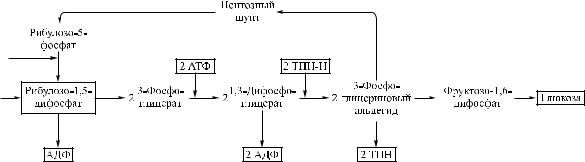
Âряде темновых реакций, происходящих в отсутствие света, ТПН·Н вызывает восстановление двуокиси углерода до углевода. Большую часть энергии, необходимой для этих реакций, доставляет АТФ. Синтез глюкозы из двуокиси углерода и воды в темновых реакциях представлен на рис. 5.3 [18].
Âэтом сложном цикле, представленном на рис. 5.3 лишь схематиче- ски, главным промежуточным продуктом служит рибулозодифосфат, который присоединяет к себе двуокись углерода и образует две молекулы
73
74
Рис. 5.3. Синтез глюкозы из двуокиси углерода и воды в темновом цикле Кальвина
фосфоглицерата. Фосфоглицерат восстанавливается при участии ТПН·Н и в результате ряда реакций превращается в глюкозу. Рибулозодифосфат образуется вновь в процессе реакций, названных пентозным шунтом. Молекула глюкозы, образовавшаяся в ряде реакций темнового цикла, является конечным продуктом и содержит значительное количество солнечной энергии. Большинство гетеротрофных микроорганизмов используют энергию, заключенную в молекуле глюкозы, постепенно расщепляя ее. Извлеченная из молекулы глюкозы энергия связывается молекулой АТФ в процессе дыхания.
5.4. Дыхательная цепь
Дыхание — это окисление органических веществ. Процессы ассимиляции пищи протекают с затратой энергии.
Потребность в энергии обеспечивается процессами энергетического обмена, сущность которых заключается в окислении органических веществ, сопровождаемом выделением энергии. Получаемые при этом продукты окисления выделяются в окружающую среду. Окисление веществ может происходить различными путями:
1. Прямое присоединение к веществу кислорода:
C6Í12Î6 + 6Î2 6ÑÎ2 + 6Í2Î + 2,8·106 êÄæ.
2. Непрямое окисление, дегидрогенирование (отнятие водорода):
ÀÍ2 + Â дегидрогеназа À + ÂÍ2 + энергия,
где А, В — различные вещества, на которые действует фермент. Окисляемое вещество или отдает водород, или присоединяет моле-
кулу воды, а затем отщепляется водород (окисление вещества происходит в этом случае за счет кислорода воды). Отнятый от окисляемого вещества водород переносится на другое вещество, которое при этом восстанавливается. Вещество, отдающее водород, называется донором,
àвещество, которому передается водород, акцептором.
3.Перенос электронов (å–) от одного вещества к другому:
Fe3+ + å– Fe2+ – å– Fe3+.
Вещество, теряющее электроны, окисляется, а присоединяющее их — восстанавливается.
75
Âживых клетках окисление органических веществ происходит чаще всего путем дегидрогенирования. Атом водорода состоит из протона (Н+) и электрона (å–), т. е. перенос водорода с одного вещества на другое включает и перенос электрона.
Перенос водорода (электрона) от окисляемого вещества к акцептору осуществляется различными оксидоредуктазами — окислительно-вос- становительными ферментами. Конечным (терминальным) акцептором водорода может быть кислород воздуха или другое вещество, способное восстанавливаться. В процессе дыхания животные и растительные организмы окисляют органические соединения до углекислого газа и воды, потребляя при этом кислород воздуха.
Способы получения энергии у микроорганизмов разнообразны. Аэробы, подобно высшим организмам, получают энергию, окисляя
органические вещества с использованием молекулярного кислорода.
Ó анаэробов окислительные процессы протекают без участия кислорода. Анаэробные микроорганизмы по отношению к кислороду подразделяются на облигатные (строгие, безусловные), для которых кислород не только не нужен, но и вреден, и факультативные (условные, не обязательные), они могут жить как при доступе воздуха, так и без него. Степень анаэробности у факультативных анаэробов различна. Одни лучше развиваются в анаэробных условиях или при ничтожно малом содержании кислорода в среде (микроаэрофилы), другие — при доступе воздуха. Известны факультативные анаэробы (некоторые дрожжи), способные
âзависимости от условий развития переключаться с анаэробного на аэробный тип получения энергии.
Âэнергетическом обмене микроорганизмы могут использовать природные органические и многие синтетические соединения, но не все из них способны ассимилировать любые вещества. Одни способны окислять небольшое число соединений, другие — использовать в энергетиче- ских целях узкоспецифические вещества субстрата.
Независимо от того, к какому типу относится тот или иной микроорганизм, биологическая сущность осуществляемых ими окислительных процессов одна — получение энергии. Поэтому эти процессы правильнее называть не дыханием, а биологическим окислением.
Аэробные микроорганизмы (грибы, многие бактерии) используют
âкачестве энергетического материала в основном глюкозу. В процессе катаболизма центральным амфиболитом считается глюкоза, которая
76
образуется при фотосинтезе и может вступать в процесс получения энергии. При полном окислении глюкозы освобождается вся потенциальная энергия, аккумулированная в ее молекуле в процессе фотосинтеза из СО2 è Í2О. Имеются три основных пути окисления глюкозы: гликолиз, пентозофосфатный è кетодезоксифосфоглюконатный (ÊÄÔÃ-ïóòü) [1].
Гликолиз, или путь Эмбдена — Мейергофа — Парнаса, встречается у прокариот и эукариот. Из молекулы глюкозы образуются две молекулы пирувата и синтезируются 2НАД·Н и 2АТФ.
Пентозофосфатный путь (гексозомонофосфатный, или путь Варбурга — Диккенса — Хорекера) распространен у растений, а у микроорганизмов играет вспомогательную роль. В результате реакций образуются пентозы для последующих синтезов: пируват, три молекулы пиридиннуклеотидов и две молекулы АТФ.
ÊÄÔÃ-ïóòü, или путь Энтнера — Дуборова, присущ только микроорганизмам и приводит к образованию из одной молекулы глюкозы двух молекул пирувата, молекулы АТФ и двух молекул восстановленного пиридиннуклеотида.
Первостепенное значение имеет разложение глюкозы в процессе гликолиза, или путь Эмбдена — Мейергофа — Парнаса.
Процесс окисления глюкозы подразделяется на две фазы: анаэробную — гликолиз и аэробную — цикл Г. Кребса, или цикл трикарбоновых кислот.
Гликолиз является первой (подготовительной) фазой процесса окисления глюкозы, в результате которого происходит расщепление шестиуглеродной молекулы глюкозы на две трехуглеродные молекулы молоч- ной кислоты или ее окисной формы — пировиноградной кислоты. Этот процесс состоит из 11 ступеней, каждая из которых катализируется своим особым ферментом. Начальные этапы окисления глюкозы проходят однотипно как у аэробов, так и у анаэробов. Суть процесса гликолиза заключается не в разделении молекулы глюкозы на части, а в последовательном извлечении из нее энергии [18].
В ходе превращения глюкозы в пировиноградную кислоту протекают следующие реакции:
1. Активирование глюкозы путем фосфорилирования (присоединение фосфорного остатка от молекулы АТФ) при участии фермента гексокиназы (фосфотрансферазы):
глюкоза + АТФ гексокиназа глюкозо-6-фосфат + АДФ.
77
2. Изомеризация — перенос фосфорной группы под действием фермента глюкозофосфатизомеразы:
глюкозо-6-фосфат глюкозофосфатизомераза фруктозо-6-фосфат.
3. Фосфорилирование молекулы фруктозо-6-фосфата при участии АТФ и фермента фосфотрансферазы (фосфофруктокиназы):
фруктозо-6-фосфат + АТФ фосфотрансфераза фруктозо-1,6-дифосфат +
+ ÀÄÔ.
4. Расщепление фруктозо-1,6-дифосфата на две фосфотриозы: фосфатдиоксиацетон è фосфоглицериновый альдегид — под действием фермента альдолазы:
фруктозо-1,6-дифосфат альдолаза фосфатдиоксиацетон +
+ фосфоглицериновый альдегид.
Оба эти вещества легко могут превращаться друг в друга.
5. Изомеризация двух молекул 3-фосфоглицеринового альдегида (фосфатдиоксиацетон под действием фермента триозофосфатизомеразы превращается в 3-фосфоглицериновый альдегид):
фосфатдиоксиацетон триозофосфатизомераза 3-фосфоглицериновый
альдегид.
6. Окисление 3-фосфоглицеринового альдегида под действием фермента дегидрогеназы, коферментом которой является НАД:
3-фосфоглицериновый альдегид + 2Н ÐÎ дегидрогеназа, 2НАД, 2ТПН 3 4
2 1,3-дифосфоглицериновая кислота + 2НАД·Н + 2ТПН·Н.
К 3-фосфоглицериновому альдегиду присоединяются две молекулы фосфорной кислоты с участием двух молекул ТПН и кофермента НАД. Молекула 3-фосфоглицеринового альдегида присоединяет фосфат, а водород переносится на кофермент НАД и ТПН, которые восстанавливаются. Освобожденная энергия при окислении 3-фосфоглицеринового альдегида сосредоточивается в макроэргической связи образующейся 1,3-дифосфоглицериновой кислоты.
7. Подзарядка АТФ.
Фосфатный остаток 1,3-дифосфоглицериновой кислоты, имеющей макроэргическую связь, при участии фермента фосфотрансферазы ïå-
78
реносится на молекулу аденозиндифосфорной кислоты, и образуется 3-фосфоглицериновая кислота, а АДФ превращается в АТФ:
1,3-дифосфоглицериновая кислота + 2АДФ |
фосфотрансфераза |
|
2 3-фосфоглицериновая кислота + 2 АТФ.
8. 3-Фосфоглицериновая кислота под влиянием фермента фосфоглицеромутазы превращается в 2-фосфоглицериновую кислоту:
2 3-фосфоглицериновая кислота — 2Р |
фосфоглицеромутаза |
|
2-фосфоглицериновая кислота.
9. Под действием фермента енолазы 2-фосфоглицериновая кислота, теряя воду, переходит в енольную форму фосфопировиноградной кислоты (енолфосфопируват):
2-фосфоглицериновая кислота 2H2O
енолаза
2 фосфоенолпировиноградная кислота + 2Н 2Î.
При этом превращении происходит перераспределение внутримолекулярной энергии, большая часть которой сосредоточивается в форме макроэргический фосфатной связи фосфоенолпировиноградной кислоты.
10. Вторая подзарядка АДФ.
Фосфоенолпировиноградная кислота дефосфорилируется, при этом остаток фосфорной кислоты при участии фермента пируваткиназы передается вместе с макроэргической связью на молекулу АДФ. В результате образуется пировиноградная кислота, а АДФ превращается в АТФ:
2 фосфоенолпировиноградная кислота + 2 АДФ пируваткиназа
2 пировиноградная кислота + 2 АТФ.
11. Восстановление пировиноградной кислоты ТПН·Н до молоч- ной кислоты под действием фермента енолазы:
2ÑÍ3 – ÑÎ – ÑÎÎÍ + 2ÒÏÍ·Í |
енолаза |
|
2ÑÍ3 – ÑÍ(ÎÍ) – ÑÎÎÍ + 2ÒÏÍ.
После расщепления глюкозы до двух молекул молочной кислоты аэробные клетки продолжают извлекать большую часть энергии в процессе дыхания, во время которого трехуглеродная молекула молочной кислоты расщепляется на одноуглеродные молекулы двуокиси углерода.
79
Эти начальные этапы окислительного превращения глюкозы до молочной кислоты проходят однотипно как у аэробов, òàê è ó анаэробов без участия кислорода.
Окисленная форма молочной кислоты — пировиноградная кислота — претерпевает более сложный ряд превращений, получивших название цикла трикарбоновых кислот èëè цикла Кребса, каждая реакция которого катализируется особой ферментативной системой. Цикл превращений пировиноградной кислоты у аэробов описан впервые Г. Кребсом в 1937 г. Это открытие является краеугольным камнем в биохимии.
Схема цикла представлена на рис. 5.4.
Цикл Кребса позволяет проследить окисление окисной формы молочной кислоты до двуокиси углерода. Однако этот цикл не объясняет, каким образом энергия, заключенная в молекуле молочной кислоты, извлекается в форме, пригодной для использования в живой клетке.
В настоящее время полагают, что в процессе цикла Кребса электроны при участии ферментов отрываются от промежуточных продуктов и передаются последовательно ряду молекул-переносчиков, объединенных под общим названием дыхательной цепи. Эта цепь ферментных молекул представляет собой конечный общий путь всех электронов, отнятых от молекул питательных веществ в процессе биологического окисления. В последнем звене этой цепи электроны соединяются с кислородом, образуя воду. Водород, отщепленный от окисляемых в цикле Кребса кислот, передается посредством коферментов (НАД и ФАД) и соответствующих дегидрогеназ по дыхательной цепи, состоящей из комплекса ферментов, к конечному акцептору — молекулярному кислороду. Водород от восстановленного НАД·Н передается на кофермент ФАД флавинового фермента, который восстанавливается в ФАД·Н. Далее с восстановленной флавиновой дегидрогеназы водород передается на цитохром цитохромной системы. Атом водорода расщепляется на ион водорода (Н+) и электрон, т. к. цитохром способен воспринимать только электроны (å–) от атомов водорода. Цитохром из окисленной формы переходит в восстановленную форму. Последний цитохром передает электроны цитохромоксидазе, восстанавливая ее кофермент. Завершающей реакцией является окисление восстановленной цитохромоксидазы молекулой кислорода. Кислород активируется за счет передачи ему электронов с цитохромоксидазы и приобретает способность соединяться с ионами водорода, в результате чего образуется вода. Освобождающаяся в процессе переноса электронов энергия затрачивается на синтез АТФ из АДФ.
80