

1. Регуляция всасывания в кишечнике.
2. Поддержание рециркуляции железа эритроцитов.
3. Регуляция количества поступающего в клетку железа.
ЭТАПЫ ОБМЕНА ЖЕЛЕЗА В ОРГАНИЗМЕ. Кратко остановимся на процессе всасывания железа, состоящем из ряда последовательных этапов:
начальный захват железа щеточной каймой клеток слизистой оболочки кишечника;
внутриклеточный транспорт его образование лабильных запасов железа в клетке;
3.освобождение железа из слизистой оболочки кишечника в кровь.
3.Задача,ОТВЕТ: а) Фактор V и фактор VIII – доменные белки, циркулирующие в крови. Фактор V синтезируется в печени, а фактор VIII – эндотелиальными клетками. Факторы Va и VIIIa служат белками-активаторами факторов VIIa и Xa. При связывании с белками-активаторами в результате конформационных изменений активность этих ферментов повышается.
б) Существует 2 пути свертывания крови –прокоагулянтный (внешний) и контактный (внутренний). Для инициации реакций внешнего пути необходимо появление тканевого фактора (фактор III) на внешней поверхности плазматической мембраны клеток, соприкасающихся с кровью. Внутренний путь начинается с активации фактора XII при его контакте с поврежденной поверхностью эндотелия сосудов и взаимной активации ферментов прекалликреина и фактора XII.

Рис. 82 Система коагуляции
В прокоагулянтном и контактном путях свертывания крови последовательное образование мембранных ферментных комплексов активируется фактора Х и образуются протромбиназы. Этапы, одинаковые для обоих путей свертывания крови, называются общим путем свертывания крови.
в) Протеин С участвует в процессе антикоагуляции. Активированный протеин С (Са) образует с белком-активатором S мембраносвязанный комплекс Са-S-Са2+. Протеин Са в составе этого комплекса гидролизует в факторах Va и VIIIa по 2 пептидные связи и инактивирует эти факторы.

1.
2,3-ДФГ снижает сродство гемоглобина к кислороду и, таким образом, повышает отдачу кислорода тканям. Если кровь израсходовала весь свой запас ДФГ, гемоглобин остается фактически насыщенным кислородом. При акклиматизации в условиях высокогорья содержание ДФГ в эритроцитах резко увеличивается. ДФГ является аллотерическимлигандом, так как связывается с гемоглобином в другом по сравнению с О2 участком. ДФГ встраивается в полость тетрамерной молекулы гемоглобина, полость образована остатками всех 4 протомеров.

2.

В данном случае используется способность молекул IRP связываться с участком IRE соответствующих мРНК для двух белков – рецептора трансферрина и ферритина. При этом в комплексе с железом IRP не активен и не присоединяется к мРНК, без железа - может присоединяться.
1. Присоединение irp к мРнк рецептора трансферрина ближе к ее 3'-концу защищает мРнк от разрушения рнКазами, действующими с 3'-конца мРнк:
при низкой концентрации железа в клетке белок IRP является активным, присоединяется к мРНК и, как следствие, мРНК рецепторов трансферрина существует дольше, образуется больше белков-рецепторов и повышается поток железа в клетки.
при высоком содержании железа в клетке белок IRP присоединяет железо, становится неактивным, с мРНК связаться не может и, соответственно, не защищает ее от разрушения. Синтез рецепторов к трансферрину не происходит, дополнительное железо клеткой не захватывается.
Метаболизм железа
После всасывания железо либо откладывается в клетках кишечника в составе ферритина, либо сразу попадает в кровоток и в комплексе с трансферрином переносится в клетки печени, костного мозга или других тканей. Нагруженный железом трансферрин (холотрансферрин) взаимодействует со своим специфическим мембранным рецептором. Трансферриновый рецептор на низком уровне экспрессируется во всех типах клеток, на высоком уровне – в активно делящихся клетках.
Наибольшее количество трансферриновых рецепторов находится на поверхности эритробластов (300 тыс - 400 тыс на одну клетку). Период полужизни комплекса трансферрин-Fe не превышает 60-90 мин и большая часть железа, транспортируемая трансферрином, поступает в костный мозг. При усиленном эритропоэзе период полужизни комплекса сокращается до 10-15 мин, в противоположной ситуации он удлиняется до нескольких часов.
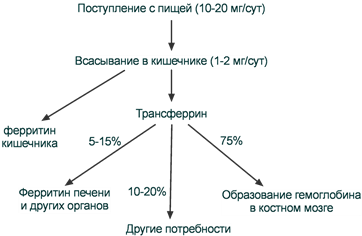
Пути поступления и использования ионов железа
Роль рецептора к трансферрину
После образования тройного комплекса (железо, апотрансферрин, рецептор) он перемещается в цитоплазму эндоцитозом и оказывается внутри эндосом. На мембране эндосомы присутствует H+-АТФаза, создающая кислый рН внутри эндосом. В результате железо диссоциирует из комплекса и составляет свободный пул.