

3.2. Слуховая сенсорная система
Слуховая сенсорная система — сенсорная система, обеспечивающая кодирование акустических стимулов и обусловливающая способность животных ориентироваться в окружающей среде посредством оценки акустических раздражителей. Периферические отделы слуховой системы представлены органами слуха и лежащими во внутреннем ухе фонорецепторами. На основе формирования сенсорных систем (слуховой и зрительной) формируется назывательная (номинативная) функция речи - ребенок ассоциирует предметы и их названия.
В качестве рецептивных полей слуховой системы могут рассматриваться части слухового пространства (англ. auditory space) или диапазоны слуховых частот. Лишь немногие исследователи трактуют слуховые рецептивные поля как определённые участки сенсорного эпителия, например, группы волосковых клеток спирального органа улитки внутреннего уха млекопитающих.
Слух — способность биологических организмов воспринимать звуки органами слуха; специальная функция слухового аппарата, возбуждаемая звуковыми колебаниями окружающей среды, например, воздуха или воды. Одно из биологических пяти чувств, называемое также акустичеcким восприятием.
Человек способен слышать звук в пределах от 16 Гц до 22 кГц при передаче колебаний по воздуху, и до 220 кГц при передаче звука по костям черепа. Эти волны имеют важное биологическое значение, например, звуковые волны в диапазоне 300—4000 Гц соответствуют человеческому голосу. Звуки выше 20 000 Гц имеют малое практическое значение, так как быстро тормозятся; колебания ниже 60 Гц воспринимаются благодаря вибрационному чувству. Диапазон частот, которые способен слышать человек, называется слуховым или звуковым диапазоном; более высокие частоты называются ультразвуком, а более низкие — инфразвуком.
Способность различать звуковые частоты сильно зависит от конкретного человека: его возраста, пола, подверженности слуховым болезням, тренированности и усталости слуха. Отдельные личности способны воспринимать звук до 22 кГц, а возможно — и выше.
Некоторые животные могут слышать звуки не слышимые человеком (ультра- или инфразвук). Летучие мыши во время полёта используют ультразвук для эхолокации. Собаки способны слышать ультразвук, на чём и основана работа беззвучных свистков. Существуют свидетельства того, что киты и слоны могут использовать инфразвук для общения.
Человек может различать несколько звуков одновременно благодаря тому, что в ушной улитке одновременно может быть несколько стоячих волн.
Удовлетворительно объяснить феномен слуха оказалось необычайно сложной задачей. Человек, представивший теорию, объяснявшую восприятие высоты и громкости звука, почти наверняка гарантировал себе Нобелевскую премию.
На сегодняшний день нет единой достоверной теории объясняющей все аспекты восприятия звука человеком. Вот некоторые из них:
Струнная теория Гельмгольца
Теория бегущей волны Бекеши
Микрофонная теория
Электро-механическая теория
Поскольку достоверная теория слуха не разработана на практике используются психоакустические модели, основанные на данных исследований проводимых на различных людях.
Опыт доказывает, что вызываемое каким-нибудь коротким звуком ощущение длится некоторое время в виде следа уже по прекращении внешнего вызвавшего его толчка. Поэтому два достаточно быстро следующих друг за другом звука дают одиночное слуховое ощущение, являющееся результатом их слияния. Но слуховые следы оказываются более кратковременными, нежели зрительные: в то время как последние сливаются уже при десятикратном повторении в секунду, для слияния слуховых ощущений требуется повторение их не менее 130 раз в секунду. Другими словами, световой след длится 1/10 с, тогда как слуховой около 1/130 секунды. Слияние слуховых ощущений имеет огромное значение в чёткости восприятия звуков и в вопросах о консонансе и диссонансе, играющих такую огромную роль в музыке.
Как бы ни возникали слуховые ощущения, мы относим их обыкновенно во внешний мир, и поэтому причину возбуждения нашего слуха мы всегда ищем в колебаниях, получаемых извне с того или другого расстояния. Эта черта в сфере слуха выражена гораздо слабее, нежели в сфере зрительных ощущений, отличающихся своей объективностью и строгой пространственной локализацией и, вероятно, приобретается также путём долгого опыта и контроля других чувств. При слуховых ощущениях способность к проецированию, объективированию и пространственной локализации не может достигнуть столь высоких степеней, как при зрительных ощущениях. Виной этому особенности строения слухового аппарата, например, такие как недостаток мышечных механизмов, лишающий его возможности точных пространственных определений. Известно то огромное значение, какое имеет мышечное чувство во всех пространственных определениях.
Наши суждения о расстоянии, на котором издаются звуки, являются весьма неточными, в особенности если глаза человека закрыты и он не видит источника звуков и окружающие предметы, по которым можно судить об «акустике окружения» на основании жизненного опыта, либо акустика окружения нетипична: так, например, в акустической безэховой камере голос человека, находящегося всего в метре от слушающего, кажется последнему в разы и даже десятки раз более удалённым. Также знакомые звуки представляются нам тем более близкими, чем они громче, и наоборот. Опыт показывает, что мы менее ошибаемся в определении расстояния шумов, нежели музыкальных тонов. Способность суждения о направлении звуков у человека весьма ограничена: не имея подвижных и удобных для собирания звуков ушных раковин, он в случаях сомнений прибегает к движениям головы и ставит её в положение, при котором звуки различаются наилучшим образом, то есть звук локализируется человеком в том направлении, с которого он слышится сильнее и «яснее».
Способность человека (и высших животных) определять направление на источник звука называется бинауральным эффектом.
Известно три механизма, при помощи которых можно различить направление звука:
Разница в средней амплитуде (исторически первый обнаруженный принцип): для частот выше 1 кГц, то есть таких, что длина звуковой волны меньше, чем размер головы слушающего, звук, достигающий ближнего уха, имеет бо́льшую интенсивность.
Разница в фазе: ветвистые нейроны способны различать фазовый сдвиг до 10-15 градусов между приходом звуковых волн в правое и левое ухо для частот в примерном диапазоне от 1 до 4 кГц (что соответствует точности в определении времени прихода в 10 мкс).
Разница в спектре: складки ушной раковины, голова и даже плечи вносят в воспринимаемый звук небольшие частотные искажения, по-разному поглощая различные гармоники, что интерпретируется мозгом как дополнительная информация о горизонтальной, а также вертикальной локализации звука.
Возможность мозга воспринимать описанные различия в звуке, слышимым правым и левым ухом, привело к созданию технологии бинауральной записи.
Описанные механизмы не работают в воде: определение направления по разности громкостей и спектра невозможно, так как звук из воды проходит практически без потерь напрямую в голову, и значит в оба уха, из-за чего громкость и спектр звука в обоих ушах при любом расположении источника звука с высокой точностью одинаковы; определение направления источника звука по фазовому сдвигу невозможно, так как из-за гораздо более высокой в воде скорости звука длина волны возрастает в несколько раз, а значит фазовый сдвиг многократно уменьшается.
Из описания приведённых механизмов понятна и причина невозможности определения расположения источников низкочастотного звука.
Слух проверяют с помощью специального устройства или компьютерной программы под названием «аудиометр».
Возможно определение ведущего уха с помощью специальных тестов. Например, в наушники подаются разные аудиосигналы (слова), а человек их фиксирует на бумаге. С какого уха больше правильно распознанных слов, то и ведущее.
Определяют и частотные характеристики слуха, что важно при постановке речи у слабослышащих детей.
Восприятие частотного диапазона 16 Гц − 22 кГц с возрастом изменяется — высокие частоты перестают восприниматься. Уменьшение диапазона слышимых частот — связано с изменениями во внутреннем ухе (улитке), и связано с развитием с возрастом нейросенсорной тугоухости.
Порог слышимости — минимальное звуковое давление, при котором звук данной частоты воспринимается ухом человека. Величину порога слышимости выражают в децибелах. За нулевой уровень принято звуковое давление 2×10−5Па на частоте 1 кГц. Порог слышимости у конкретного человека зависит от индивидуальных свойств, возраста, физиологического состояния.
Порог болевого ощущения слуховой — величина звукового давления, при котором в слуховом органе возникают боли (что связано, в частности, с достижением предела растяжимости барабанной перепонки). Превышение данного порога приводит к акустической травме. Болевое ощущение определяет границу динамического диапазона слышимости человека, который в среднем составляет 140 дБ для тонального сигнала и 120 дБ для шумов со сплошным спектром.
Преддверно-улитковый нерв
Преддверно-улитковый нерв (лат. nervus vestibulocochlearis) — нерв специальной чувствительности, отвечающий за передачу слуховых импульсов и импульсов исходящих из вестибулярного отдела внутреннего уха. Состоит из двух разных по функции корешков: вестибулярного корешка (лат. radix vestibularis), несущего импульсы от статического аппарата, представленного полукружными протоками вестибулярного лабиринта и улиткового корешка (лат. radix cochlearis) проводящего слуховые импульсы от спирального органа улиткового лабиринта (рис. 51).
На нижней поверхности мозга он показывается ниже лицевого нерва (лат. n.facialis), кнаружи от оливы продолговатого мозга.
Периферические волокна (дендриты) radix cochleare берут начало от ганглия улитки (лат. ganglion cochleare) и заканчиваются в спиральном органе, являющемся воспринимающим прибором слухового пути.
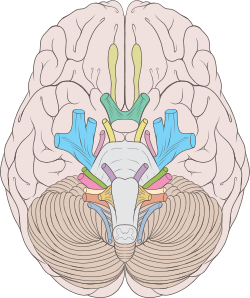
Рис. 51. Схема мозга, ствола мозга и черепно-мозговых нервов (преддверно-улитковый нерв отмечен красным цветом)
Центральные отростки (аксоны) клеток ганглия улитки образуют radix cochleare, который выходит из пирамиды височной кости через внутреннее слуховое отверстие и входит в вещество мозга. Заканчивается в заднем и переднем улитковых ядрах.
Вестибулярный корешок начинается от вестибулярного ганглия (лат. ganglion vestibulare) залегающего в расщелине внутреннего слухового прохода. Вестибулярный ганглий подразделяют на две части: верхнюю и нижнюю.
Периферические отростки (дендриты) клеток ganglion vestibulare подходят к рецепторным клеткам сферического мешочка, эллиптического мешочка и полукружным протокам. Центральные отростки (аксоны) входят в состав вестибулярного корешка, подходят к вестибулярным ядрам вестибулярного поля ромбовидной ямки (лат. fossa rhomboidea).
Слуховая система состоит из наружного, среднего и внутреннего уха. Здесь обсуждается только внутреннее ухо; состоящее из улитки (лат. cochlea), содержащей кортиев орган, спиральный орган (лат. organum spirale), и слуховой нерв. Поступающие из наружного уха звуковые волны в кортиевом органе трансформируются в нервные импульсы. Кроме воздушной проводимости имеется ещё и костная проводимость (передача звуковых колебаний через кости черепа). Из кортиева узла идут постганглионарные волокна спирального ганглия, которые направляются в этот узел и переключаются в нём образуя слуховой нерв. Слуховой нерв в свою очередь присоединяется к вестибулярному на своём пути через внутреннее слуховое отверстие височной кости. В области мостомозжечкового угла оба нерва вступают в ствол мозга непосредственно позади нижней мозжечковой ножки (лат. pedunculus cerebellaris inferior). В стволе мозга находятся вторые нейроны слухового нерва представленные передним и задним улитковыми ядрами (лат. nuclei cochleares ventralis et dorsalis), которые занимают наиболее латеральное положение вестибулярного поля ромбовидной ямки.
Аксоны, берущие начало от переднего улиткового ядра, большей частью переходят на противоположную сторону в виде «трапециевидных» волокон и участвуют в формировании трапециевидного тела, находящегося на границе между основанием и покрышкой варолиева моста. Аксоны, берущее начало из заднего улиткового ядра идут дорсально от нижней мозжечковой ножки на противоположную сторону, частично в составе мозговых полосок четвёртого желудочка (лат. striae medullares ventriculi quarti), частично в составе ретикулярной формации.
Перекрещенные волокна передают импульсы в ядро трапециевидного тела, верхнее ядро оливы, ядру латеральной петли или ретикулярную формацию. Волокна, не подвергшиеся перекресту, в основном заканчиваются в верхних оливах той же стороны. Таким образом, в верхних оливах и ядрах трапециевидного тела располагаются тела третьих нейронов слуховых путей. Их Аксоны формируют латеральную или слуховую петлю, состоящую из перекрещенных и неперекрещенных слуховых путей, которая поднимается вверх и достигает подкорковых слуховых центров — медиальных коленчатых тел и нижних бугорков четверохолмия.
Из клеток подкорковых слуховых центров берут начало последние слуховые аксоны, которые проходят через заднюю ножку внутренней капсулы и corona radiata, заканчиваясь в височной доле коры головного мозга (задний отдел верхней височной извилины и поперечные извилины Гершля, находящиеся в глубине сильвиевой борозды).
Первичное корковое поле окружено вторичными проекционными полями, в которых происходит анализ, идентификация и сравнение слуховых стимулов. Они также интерпретируются и узнаются в качестве шумов, тонов, мелодий, гласных и согласных звуков, слов и предложений, иными словами, символов речи. В случае поражения указанных корковых областей в доминантном полушарии утрачивается способность узнавать звуки и понимать речь (сенсорная афазия).
На пути от Кортиева органа до коры волокна слухового пути совершают 4-6 переключений (в ядре верхней оливы, нейронах ретикулярной формации, ядре латеральной петли, нижних холмах четверохолмия, медиальных коленчатых телах). В этих точках они отдают коллатерали, являющиеся частью рефлекторных дуг. Некоторые коллатерали связаны с мозжечком. Другие проходят по медиальному продольному пучку к ядрам, которые иннервируют мышцы глаз, и участвуют в организации содружественного поворота глаз в направлении звука. Часть волокон идёт через нижние и верхние холмики крыши среднего мозга к претектальному ядру и от него в составе тектобульбарного пути к ядрам различных черепных нервов, в том числе — к ядру лицевого нерва (для подстройки тонуса стременной мышцы (лат. m.stapedius)), а также к двигательным клеткам передних рогов шейного отдела спинного мозга. Последняя связь обеспечивает поворот головы в сторону источника звука или от него. Коллатерали, посылающие импульсы в восходящую активирующую систему ретикулярной формации, способствуют организации процесса пробуждения. Некоторые импульсы спускаются в составе латеральной петли к вставочным нейронам, оказывающим регулирующее, предположительно, частично ингибирующее влияние на тонус базальной мембраны. Считают, что эти нейроны обеспечивают способность уха сосредотачиваться на определённых частотах звука путём одновременного подавления соседних частот.
Система равновесия
Рецепторы вестибулярного анализатора располагаются в полукружных каналах и в отолитовом аппарате внутреннего уха. Отсюда импульсы следуют по дендритах первых нейронов вестибулярных путей к вестибулярному ганглию Скарне (лат. ganglion vestibulare), лежащему во внутреннем слуховом проходе. В нём располагаются тела первых чувствительных нейронов. Отсюда импульсы следуют по аксонам тех же клеток, проходящих в составе общего ствола VIII нерва. Входя в вещество мозга центральные отростки ганглия Скарпе следуют к вестибулярным ядрам, которые располагаются в проекции вестибулярного поля ромбовидной ямки на границе с мостом и продолговатым мозгом.
Комплекс вестибулярных ядер включает:
Верхнее вестибулярное ядро (ядро Бехтерева)
Латеральное вестибулярное ядро (ядро Дейтерса)
Медиальное вестибулярное ядро (ядро Швальбе)
Нижнее вестибулярное ядро (ядро Роллера)
Волокна вестибулярного нерва разделяются перед тем, как подойти к определённым клеточным группам вестибулярных ядер, где начинаются вторые нейроны. Некоторые его волокна передают импульсы непосредственно, без переключений в мозжечок, причём в его самую старую онтогенетическую часть — archicerebellum. Эфферентные импульсы от nucleus fastigii (archicerebellum) возвращаются к вестибулярным ядрам и затем по вестибулярному нерву к волосковым клеткам лабиринта, оказывая регулирующее, преимущественно ингибирующее влияние.
Archicerebellum получает также и вторичные волокна от вестибулярных ядер. Он посылает эфферентные импульсы обратно в комплекс вестибулярных ядер, а также в спинной мозг к двигательным нейронам по мозжечково-ретикулярным и ретикуло-спинномозговым связям. В латеральном вестибулярном ядре (ядре Дейтерса) начинается важный латеральный вестибулоспинальный путь. Он спускается ипсилатерально в переднем канатике к γ- и α-мотонейронам спинного мозга, достигая крестцовых сегментов. Этот путь оказывает облегчающее влияние на экстензорные рефлексы и поддерживает мышечный тонус достаточно высоким для поддержания равновесия.
Волокна от медиального вестибулярного ядра (ядра Швальбе) присоединяются с каждой стороны к медиальному продольному пучку, связываются с двигательными клетками передних рогов шейных сегментов спинного мозга и спускаются в виде медиального вестибулоспинального пути к ростральной (верхней) части грудного отдела спинного мозга. Эти волокна располагаются вблизи передней срединной борозды шейного отдела спинного мозга. Они образуют fasciculus sulcomarginalis, который спускается вниз и оканчивается в ростральной части грудного отдела спинного мозга. Эти волокна влияют на тонус мышц шеи в соответствии с различными положениями головы. Возможно также, что они принимают участие в рефлекторных дугах, способствующих поддержанию равновесия путём осуществления начальных компенсаторных движений руками.
Все вестибулярные ядра связаны с ядрами глазодвигательных нервов посредством медиального продольного пучка. Благодаря вестибуло-окуломоторным связям достигается содружественность движений глазных яблок и фиксации взора при изменении положения головы. Нарушение проводимости импульса по ним ведёт к возникновению вестибулярного нистагма. Показано, что некоторые волокна контактируют с интерстициальным ядром Кахаля и ядром Даршкевича и продолжаются до зрительного бугра.
Часть аксонов вестибулярных ядер входят в контакт с образованиями вегетативной нервной системы и, в частности с задним ядром блуждающего нерва и с ядрами гипоталамической области. Наличие этих связей объясняет появление при патологии вестибулярного анализатора выраженных вегетативных реакций в виде тошноты, рвоты, побледнения или покраснения кожных покровов, потливости, усиления перистальтики кишечника, снижения кровяного давления, брадикардии, гипергликемии и т. д.
Клиника поражения
Клинически выделяют 2 основных формы снижения слуха: глухота среднего уха, или кондуктивная глухота (связанная с нарушением звукопроведения) и глухота внутреннего уха, или невральная глухота.
Кондуктивная глухота вызывается патологическими процессами в наружном слуховом проходе или, что бывает чаще, в среднем ухе. При этом во внутреннее ухо и, следовательно, в Кортиев орган не проводятся никакие звуковые волны либо проводятся только некоторые из них. Причинами кондуктивной глухоты могут быть средний отит, отосклероз, опухоли.
При раздражении патологическим процессом периферических отделов слухового анализатора обычно возникает шум в ухе на стороне раздражения. Среди причин, обусловливающих шум в ухе, может быть воспалительный процесс, а также раздражение волокон кохлеарной порции VIII нерва, растущей из его шванновcкой оболочки опухолью — невриномой VIII нерва. Поражение улитки или ствола VIII нерва, а также его ядер его варолиевом мосту может привести к снижению слуха или глухоте на стороне патологического очага.
Если патологический процесс локализуется в области мостомозжечкового угла, то наблюдаются не только снижение слуха на стороне поражения, но и нарушении функции вестибулярного, тройничного и лицевого нервов. Кроме этого, при росте невриномы слухового нерва в сторону ствола мозга и мозжечка может сопровождаться развитием альтернирующего синдрома и присоединением мозжечковых симптомов на стороне поражения.
При локализации процесса на уровне пластинки покрышки наблюдается снижение слуха в обоих ушах с быстрым прогрессированием и развитием полной глухоты.
В связи с тем, что в латеральной петле идут как перекрещенные так и неперекрещенные волокна, то прерывание одной латеральной (слуховой) петли не вызывает односторонней глухоты. Скорее наблюдается небольшое снижение слуха с противоположной стороны (гипоакузия) и некоторое нарушение узнавания направления звука.
Если поражены слуховые пути на уровне таламуса, то у больных изменяется ощущение тональности, звуки отдаляются или приближаются. Характерными являются акустические расстройства в виде гиперпатии. Все звуки воспринимаются как очень громкие. Шум, сильные звуки вызывают болевые ощущения.
Корковые очаги в области левой (доминантного полушария) височной доли обуславливают слуховые галлюцинации. Может возникать слуховая агнозия. Если поражена правая височная доля, наблюдается слуховая гиперпатия, как и при поражении таламуса. Если патологический очаг раздражает корковый конец слухового анализатора, возникают слуховые галлюцинации, которые в таких случаях могут быть предвестником общего судорожного эпилептического припадка. Тогда говорят о наличии у больного припадков эпилепсии со слуховой аурой.
Нистагм — непроизвольные колебательные движения глаз. Так как вестибулярный нерв влияет на положение глазных яблок таким образом, что обеспечивается зрительная ориентация в пространстве при любых положениях головы. Таким образом, при каких-либо поражениях вестибулярной системы нарушается положение глазных яблок и возникает явление нистагма. О поражении вестибулярной системы свидетельствует наличие в нистагме, как быстрого, так и медленного компонента. Медленный компонент является действительным сигналом поражения, в то время как быстрый компонент обусловлен лишь толчкообразным, рефлекторным возвращением глаз в исходное положение. Принято обозначать направление нистагма в соответствии с его быстрым компонентом.
Топическая диагностика поражения разных уровней вестибулярного анализатора:
Поражение рецепторов вестибулярного анализатора в лабиринте и внутреннем ухе характеризуется системной обморочностью, горизонтальным нистагмом и снижением слуха.
Если поражён n.vestibularis, возникает системная обморочность вращающегося характера, направленная в сторону быстрого компонента нистагма. Обморочность возникает и зависит от изменения положения головы. Есть нистагм горизонтально-ротаторный, мелко- и среднеразмашистый. В позе Ромберга больной падает в сторону поражения (в сторону медленного компонента нистагма). Часто одновременно с поражением n.vestibularis отмечается поражение n.cochlearis.
Вестибулярные расстройства при наличии очагов в стволе мозга зависят от уровня поражения. Если поражено ядро Роллера (нижнее вестибулярное ядро), наблюдается ротаторный нистагм в сторону очага. При поражении ядер Швальбе и Дейтерса, наблюдается горизонтальный нистагм в одну или две стороны. Выраженность нистагма изменяется в зависимости от положения головы. Если поражено ядро Бехтерева наблюдается вертикальный нистагм. Одновременно при наличии поражения ядер ствола появляются вестибулярная атаксия и латеропульсия (отклонение во время движения в сторону).
Очаг поражения в зоне пластинки покрышки характеризуется конвергирующим нистагмом. Быстрый компонент нистагма обоих глаз направлен к середине. Выражены глазодвигательные расстройства.
При наличии поражения корковых зон (лобно-височная, теменная области) вестибулярные нарушения проявляются ощущением нестойкости, падения, колебания пола и т. д.
Периферическое поражение (лабиринтов или вестибулярных нервов) может быть вызвано следующими причинами: лабиринтит, синдром Меньера, перилимфатическая фистула, травма лабиринта (перелом пирамиды височной кости), лабиринтная апоплексия, вертебробазилярная недостаточность, токсическое поражение лабиринта стрептомицином или другими лекарствами, невринома во внутреннем слуховом проходе. Также была установлена связь между возникновением Меньеровских атак и тесным прилеганием сосуда к проксимальной немиелинизированной части корешка преддверно-улиткового нерва нерва. Оперативное перемещение сосуда приводило к исчезновению приступа болезни.
Центральное поражение может быть вызвано циркуляторными нарушениями (размягчением, кровотечением) в вертебробазилярном бассейне, рассеянным склерозом, сифилисом, опухолью или другими заболеваниями.
Методика исследования
Проверяя слух, следует иметь в виду, что при нормальном слухе человек слышит шепотную речь на расстоянии 5-6 м.
Выпадение функции n.cochlearis само собой понятно вызывает тугоухость (гипоакузию) или глухоту. Но так как эти расстройства могут наступить также и при поражениях аппарата, передающего звук, то есть среднего и наружного уха (область отоларинголога), то задачей невролога является в первую очередь определение расположения патологического процесса.
Двумя главными признаками нервной тугоухости являются понижение или отсутствие слуховой проводимости костей черепа и частичные выпадения в восприятии ряда тонов. Следует отметить, что при поражении среднего и наружного уха увеличивается время костной слуховой проводимости, что можно объяснить снижением порога чувствительности клеток кортиева органа, в который не проходят звуковые колебания из среднего уха.
Для определения костной слуховой проводимости используют следующие тесты: тест Швабаха, Вебера и Ринне.
Тест Швабаха — камертон помещается на сосцевидный отросток. При патологии внутреннего уха и n.vestibularis время костной проводимости уменьшено или равно 0. При поражении среднего уха время костной проводимости увеличивается.
Тест Ринне — предоставляет информацию о том, проводится ли звук лучше через кость или через воздух. Вибрирующий камертон ставят на сосцевидный отросток. Когда больной перестаёт его слышать, камертон помещают перед ухом исследуемого, чтобы определить, слышен ли тон камертона в этом положении. Камертон слышен, если ухо пациента здорово — позитивная проба Ринне. Если же имеется патология среднего уха, то больной слышит тон камертона через кость дольше, чем через воздух — негативная проба Ринне.
Тест Вебера — вибрирующий камертон помещают на середину темени больного. Если снижение слуха обусловлено нарушением проведения звука, больной будет слышать камертон лучше на поражённой стороне. При поражении внутреннего уха камертон лучше слышен на здоровой стороне.
В случае исследования с применением аудиометров потеря слуха в области низких частот указывает на патологию среднего уха, а потеря слуха в области высоких частот предполагает невральный генез тугоухости.
Заболевания среднего и наружного уха относятся к сфере отоларингологии. Объективные и субъективные симптомы поражения улиткового нерва и его путей должны оцениваться неврологом.
Симптомы поражения n.cochlearis могут быть вызваны невриномой слухового нерва. В этих случаях раздражение улитковых волокон в начальных стадиях ведут к появлению ушного шума в качестве первого симптома. Поражение прогрессирует очень медленно, так что нарастающее снижение слуха и нарушение определения направления звука часто ускользают от внимания больного. Обычно больные с невриномами VIII нерва обращаются к врачу, когда опухоль вырастет настолько, что начнёт повреждать соседние структуры (вестибулярный нерв, мозжечок, лицевой нерв, тройничный нерв) — синдром мостомозжечкового угла и вызовет повышение внутричерепного давления, головную боль, тошноту и рвоту.
Внезапная потеря слуха может возникать при вирусной инфекции и дисциркуляторных расстройствах, таких как вертебробазилярная недостаточность.
Другими причинами поражения Кортиева органа и n.cochlearis являются менингит, аневризмы сосудов, перилимфатическая фистула, передозировка определённых лекарственных препаратов (стрептомицина, хинина, аспирина) и сверхмощный внезапный шум (взрыв).
Центральные пути в стволе мозга страдают при сосудистых заболеваниях вследствие недостаточности кровообращения, при воспалительных процессах и опухолях. В результате возникает гипоакузия. Только двустороннее прерывание слуховых путей ведёт к двухсторонней глухоте.
Исследуют движение больного с раскрытыми и закрытыми глазами, ставя их в позу Ромберга. Неустойчивость в позе Ромберга усиливается во время вращения головы в стороны.
Тест Миттельнаера — больного просят делать «шаг на месте». Постепенно больной поворачивается в сторону очага раздражения в вестибулярном анализаторе.
Также большую помощь в исследовании системы равновесия является выявление нистагма при исследовании глазодвигательных нервов. Правильная интерпретация выявленного нистагма позволяет провести топическую диагностику поражения вестибулярной системы.
Евстахиева труба
Евстахиева труба (или слуховая труба; лат. tuba Eustachii, tuba auditiva) — канал, сообщающий полость среднего уха с глоткой. Назван в честь Бартоломео Эустахио — итальянского врача и анатома XVI века.
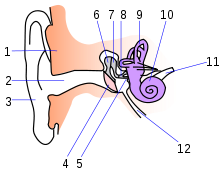
Рис. 52. Строение уха: евстахиева труба — 12
Впервые появляется у амфибий, хотя у бесхвостых и некоторых других амфибий и амфисбен между рептилиями её нет, как нет и среднего уха вообще. Нет Евстахиевой трубы у змей. Оба эти случая представляют, вероятно, результат упрощения.
Нередко отверстия обеих труб слиты в одно (безъязычные амфибии и некоторые рептилии). Морфологически Евстахиева труба представляет часть жаберной щели, а физиологически служит для уравновешивания разницы атмосферного давления извне и в полости среднего уха.
Иннервируется веточкой барабанного нерва, который в свою очередь является ветвью языкоглоточного. Воспаление слуховой трубы называется тубоотит (евстахиит).