

1. Активный транспорт требует гидролиза аденозинтрифосфата (атф).
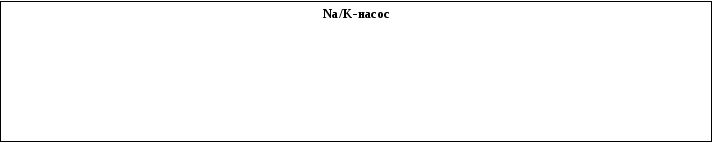
• Первично активный транспорт идет через мембранные белки, которые как только соединяются с АТФ, гидролизуются для движения растворенного вещества. Наиболее важный процесс активного транспорта связан с работой Na/K-насоса, непрерывно откачивающего Na+ из клетки в обмен на K+ с использованием энергии расщепления АТФ. На внутренней стороне мембраны 3 иона Na+ соединяется с молекулой белка-переносчика. Образовавшийся комплекс конформируется и ионы Na+ оказываются на наружной стороне мембраны. Комплекс распадается, а освободившийся переносчик соединяется с 2 ионами K+ и транспортирует их внутрь клетки. Ионы K+ освобождаются в цитоплазму, и цикл повторяется (до 200 раз в секунду). Таким образом, соотношение числа переносимых за один цикл работы фермента ионов Na+ и K+, и, соответственно, электрических зарядов, равно 3/2. Следовательно, ионный насос не только изменяет концентрацию, но и является электрогенным – создает поток положительных зарядов из клетки.
• Вторично активный транспорт соединяет транспорт двух или нескольких растворенных веществ вместе. Во вторично активном транспорте энергия используется для разработки благоприятной электрохимической движущей силы для одного растворенного вещества, которое затем используется для возможности перевозки других растворенных веществ (например, направленный внутрь Na+ градиент используется для поглощения глюкозы из кишечника). Котранспорт (симпорт) объединяет движение двух или более растворов в одном направлении. Пример Na+-переносчики включают котранспорт Na+ / глюкозы в кишечнике. Обменник (антипорт) объединяет движение двух растворенных веществ в противоположном направлении. Пример Na+-переносчики включают Na+/Ca2+ и Na+ /H+ обмен, которые являются важными для поддержания низких внутриклеточных [Ca2+] и [H+], соответственно.
Везикулярный транспорт. Движение макромолекул происходит в ограниченных мембраной пузырьках; макромолекулы проникают в клетки путем эндоцитоза и выходят из клеток путем экзоцитоза. Эндоцитоз можно описать как прием внеклеточного материала для формирования эндоцитозных пузырьков внутри клетки. Есть три типа эндоцитоза: 1) Пиноцитоз это употребление малых частиц из ECF, он происходит в большинстве клеток. 2) Фагоцитоз это поглощение крупных частиц (например, микроорганизмов), которое происходит в специализированных иммунных клетках. 3) Рецептор-опосредованный эндоцитоз это поглощение специфических молекул, он происходит на специализированных областях мембраны клатриновых ямах (например, поглощение холестерина связан с липопротеинами низкой плотности). Экзоцитоз это экспорт растворимых белков в межклеточное пространство. Такие белки синтезируются в клетке и упаковывается во внутриклеточные везикулы. Когда пузырьки сливаются с плазматической мембраной, растворимые белки выделяются и мембраны пузырьков включаются в плазматическую мембрану.
1.2 «Животное электричество». Опыты Гальвани и Матеучи
В конце XVIII в. (1786) профессор анатомии Болонского университета Луиджи Гальвани провел ряд опытов, положивших начало целенаправленным исследованиям биоэлектрических явлений. В первом опыте, подвешивая с помощью медного крючка на железной решетке препарат задних лапок лягушек со снятой кожей, ученый обнаружил, что всякий раз, когда мышцы касались решетки, они отчетливо сокращались. Л. Гальвани высказал предположение о том, что сокращение мышц является следствием воздействия на них электричества, источником которого выступают «животные ткани» — мышцы и нервы.
Однако другой итальянский исследователь — физик и физиолог Вольта — оспорил это заключение. По его мнению, причиной сокращения мышц был электрический ток, возникающий в области контакта двух разнородных металлов: меди и железа (гальваническая пара) — с тканями лягушки. С целью проверки своей гипотезы Л. Гальвани поставил второй опыт, в котором нерв нервно-мышечного препарата набрасывался на мышцу стеклянным крючком так, чтобы он касался поврежденного и неповрежденного ее участков. В этом случае мышца также сокращалась. Во втором опыте были получены абсолютные доказательства существования «животного электричества».
Окончательное доказательство существования электрических явлений в живых тканях было получено в опыте Матеуччи, в котором один нервно-мышечный препарат возбуждался током, а биотоки сокращающейся мышцы раздражали нерв второго нервно-мышечного препарата.