

7 Подвижность..Наиболее отчетливое движение бактерий видно в препарате «висячая капля» Подвижность клетки оживлено, передвигаются в поле зрения микроскопа в разных направлениях . В препарате «висячая капля» подвижность устанавливают только для форм, имеющих жгутики или способны передвигаться благодаря сокращению тела.
Для приготовления «висячей капли» исследуемый матерная наносят на середину обезжиренного покровного стекла Затем берут предметное стекло с лункой, вокруг лунки наносят тонкий слой вазелина и стекло, повернув лункой вниз прикладывают к покровному так . чтобы капля находилась в центре углубления. Предметное и покровное стекло переворачивают и капля оказывается герметично закрытой во влажной камере и защищенной от высыхания. Для того , чтобы стенки образовавшейся камеры не запотели, предметное стекло и лунку слегка увлажняют. Для исследования используют молодые (6-12-18 часовые, но не старые односуточные) бульонные культуры.
Препарат «раздавленная капля»:
На предметное стекло наносит каплю физиологического раствора в которую стерильной бактериологической петлей наносят микробную культуру в растирают ее в суспензию, после чего накрывают ее покровным стеклом так . чтобы в жидкости не образовалось пузырьков воздуха Излишки выступленной жидкости из-под стекла удаляют полосками фильтровальной бумаги. Препарат рассматривают с объективом на х 8, и на х 40.
При наблюдении под микроскопом можно заметить активное движение бактерий, которые перемещаются в различных направлениях с разной скоростью.
8. Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Цитоплазматическая мембрана Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы). Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде. Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки -. рода Pseudomonas. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.- Spirillum
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, Споры кислотоустойчивы, поэтому окрашиваются по по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет. Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
9. если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. Окраска по Граму позволяет отличить бактерии , чья толстая клеточная стенка практически полностью состоит из пептидогликана ( грамположительные ), от бактерий, чья клеточная стенка помимо тонкого слоя пептидогликана имеет наружную мембрану, состоящую из липопротеидов и липополисахаридов ( грамотрицательных ). Основный краситель (например, кристаллический фиолетовый) прочно фиксируется в стенке грамположительных бактерий, придавая им иссиня-черный цвет, и легко вымывается спиртом (или ацетоном) из стенки грамотрицательных бактерий, после чего они докрашиваются контрастным красителем (например, сафранином) в красный цвет. Микроскопию мазков кала, окрашенных по Граму, используют как отборочное исследование. При обнаружении нейтрофилов приступают к бактериологическому исследованию и определению токсинов, вырабатываемых Clostridium difficile .
Окраску по Граму используют также для выявления бактерий и лейкоцитов в СМЖ, синовиальной, плевральной и перитонеальной жидкости.
Грамположительные бактерии составляют меньшую часть тех бактерий, которые изучает меди-цинская микробиология. Ниже приводятся основные (этот перечень будет позже добавлен неспорооб-разующими анаэробами). 1. Грамположительными являются большинство кокков (кроме нейссерий): стафилококки, стрепто-кокки и пневмококки, энтерококки. 2. Среди палочек грамположительными являются листерии, бактерии актиномицетного ряда (актино-мицеты, микобактерии, коринебактерии), спорообразующие палочки (бациллы и клостридии). Е. Большинство бактерий, имеющих медицинское значение, грамотрицательные. Лишь по отношению к микоплазмам не корректно говорить об их грамотрицательности (хотя по Граму они окрашивались бы в розовый цвет – если бы их так окрашивали, так как на практике этот метод в изучении микоплазм не применяется). Дело в том, что грамоложительные и грамотрицательные бактерии отлича-ются друг от друга типом клеточной стенки (прежде всего – количеством содержащегося в ней пепти-догликана), а у микоплазм клеточной стенки с содержанием пептидогликана нет. 1. Из кокков грамотрицательные – нейссерии. 2. Грамотрицательными являются большинство палочек (собственно все, за исключением перечисленных выше грамположительных). 3. Клеточную стенку грамотрицательного типа имеют также спирохеты
10. Универсальным методом окраски микроорганизмов является окраска по Романовскому-Гимзе (смесью азура, эозина и метиленового синего). При окрашивании простейших их цитоплазма приобретает голубой цвет, а ядра - красно-фиолетовый. Этот метод используют также при исследовании риккетсий, хламидий, спирохет, форменных элементов крови Метод Циля-Нильсена предназначен для дифференциации кислотоустойчивых бактерий (возбудителей туберкулеза и лепры) от некислотоустойчивых.1.Мазок окрашивают карболовым фуксином Циля (основной краситель) при нагревании 3-5 мин.
2.Обесцвечивают раствором серной кислоты (дифференцирующее вещество) в течение 1-2 мин.
3.Промывают водой.
4.Докрашивают 3-5 мин метиленовым синим (дополнительный краситель).
Клеточная стенка кислотоустойчивых бактерий отличается высоким содержанием липидов. Они с трудом окрашиваются, но затем удерживают основной краситель при обесцвечивании кислотой. Некислотоустойчивые бактерии легко окрашиваются, а затем легко обесцвечиваются кислотой и окрашиваются дополнительным красителем.
Метод Ожешко сходен с методом Циля-Нильсена, но отличается использованием раствора соляной кислоты в качестве протравы, разрыхляющей оболочку споры, которая плохо воспринимает красители. После протравы соляной кислотой при нагревании в течение 2-3 мин мазок фиксируется и окрашивается по методу Циля-Нильсена.
1.Фиксированный мазок окрашивают уксуснокислой синей 4 мин, затем сливают краску.
2.Промывают водой и наливают раствор Люголя на 20-30 сек.
3.Не промывая водой, окрашивают везувином 1-3 мин.
4.Промывают водой, высушивают.
NB! Тела бактерий окрашиваются в нежный светло-коричневый цвет, зерна волютина - в темно-синий, почти черный цвет.
Метод Нейссера используется для выявления зерен волютина. Мазок окрашивается уксуснокислым метиленовым синим 2-3 минуты, при этом происходит химическое взаимодействие красителя и волютина. Зерна волютина окрашиваются в черный цвет. При промывке водой тело клетки обесцвечивается и затем в течение 1 мин докрашивается везувином в желто-коричневый цвет. Метод Бурри-Гинса используется для окраски капсульных бактерий и основан на том, что капсула не воспринимает красители. Капсулу выявляют негативным контрастированием фона по Бурри. Для этого черную тушь смешивают в культурой и высушивают. После этого проводят фиксацию в пламени горелки, окрашивают тела микробных клеток по Гинсу - водным фуксином в течение 1 минуты и промывают водой 5-10 секунд.
NB! В результате на темном фоне хорошо видна бесцветная капсула и красные тела микробов.
11. Морфология риккетсий и хламидий. Риккетсии и хламидии входят в отдел Gracilicutes и составляют соответственно роды Rickettsia,Chlamidia и Chlamydophila. Относятся к порядку Rickettsiаles. Он включает в себя 3 семейства: Rickettsiаceae, Bartonellaceae, Anaplasmataceae. Они являются энергетическими облигатными внутриклеточными паразитами. У них отсутствует система регенерации АТФ. Факторы патогенности.Имеют токсины: эндотоксин (ЛПС); другой токсический фактор – термолабильный белок, который находится в капсулоподобном слое, способен под действием формалина приобретать свойства анатоксина. Размножаются в основном внутриклеточно, чаще в цитоплазме. Они размножаются: в желточном мешке куриного эмбриона(оплодотворенные куриные яйца инкубируют около семи суток в термостате, после чего, проделав небольшое отверстие в скорлупе, вводят в них содержащий риккетсии материал. Примерно через восемь суток после заражения эмбрионы погибают в результате размножения и накопления риккетсий. Содержимое желточного мешка зараженных эмбрионов собирают, риккетсии очищают и концентрируют, а затем инактивируют с помощью химических реагентов или ультрафиолетового облучения. Такую суспензию инактивированных риккетсий используют в качестве вакцины против сыпного тифа или других риккетсиозных заболеваний.), в организме экспериментальных животных, в тканевых культурах.
Имеют все структуры, присущие прокариотам: клеточную стенку (в ней содержится мурамовая кислота), нуклеоид, рибосомы. Спор, жгутиков, капсул не имеют.
Грамотрицательны, окрашиваются по Романовскому-Гимзе в лиловый цвет, по Здродовскому (аналог метода Циль-Нильсена) – в красный. Риккетсии полиморфны, т. е. имеют различные морфологические формы: кокковидные (0,5 мкм); палочковидные (1,5 мкм); бациллярные (2-4 мкм); нитевидные (10-40 мкм).
Размножаются риккетсии простым делением, а нитевидные формы – дроблением. Вызывают сыпной тиф идругие риккетсиозы. обычно паразитирующие внутри эпиталиальных клеток кишечного тракта насекомых и клещей. Лишь один вид риккетсий Rochalimaea quintana, вызывающий окопную лихорадку, может расти вне клеток в кишечнике вши, а также в бесклеточной питательной среде. П. Ф. Здродовский разделяет риккетсиозы человека на пять групп: I — группа сыпного тифа, включающая эпидемический(Rickettsia prowazekii и эндемический сыпной тиф; Источник инфекции – больной человек.вши. Имеют два антигена: 1) группоспецифический (обладает иммуногенными свойствами); 2) корпускулярный, видоспецифический (имеется только у данного вида). Специфическая профилактика: живая сыпнотифозная вакцина. Осложнения- тромбофлебиты, эндартерииты, тромбоэмболия легочных артерий, кровоизлияние в мозг, миокардиты. Преимущественная локализация в центральной нервной системе приводит к осложнениям в виде психоза, При сыпном тифе характерным является умеренный нейтрофильный лейкоцитоз с палочкоядерным сдвигом, эозинопения и лимфопения, умеренное повышение СОЭ. терапию антибиотиками тетрациклиновой группы. Эндемический -Возбудитель – R. typhi. Источник инфекции – крысиные блохи, вши, гамазовые клещи. Пути заражения – трансмиссивный, воздушно-капельный. имеют видоспецифический антиген II — группа клещевой пятнистой лихорадки, куда относятся: 1) пятнистая лихорадка Скалистых гор Америки-— Rickettsia rickettsii( клещи) первичный аффект на месте укуса клеща не образуется. Риккетсии по лимфатическим путям проникают в кровь, паразитируют не только в эндотелии сосудов, но и в мезотелии, в мышечных волокнах. Инкубационный период длится от 3 до 14 дней При тяжелых формах болезни рано проявляется тромбогеморрагический синдром (носовые кровотечения, рвота «кофейной гущей», геморрагическая сыпь, кровоподтеки на месте инъекций и др. 2) марсельская, или средиземноморская, лихорадка; 3) североавстралийский клещевой сыпной тиф; 4) клещевой риккетсиоз, или клещевой сыпной тиф Средней Азии; 5) осповидный, или везикулярный, риккетсиоз; III — группа краснотелково-клещевой лихорадки, или группа цуцугамуши, куда входит японская речная лихорадка цуцугамуши; IV — группа пневмотропных риккетсиозов, или группа Ку- лихорадк; Возбудитель –Coxiella burneti. Источник инфекции – домашний скот. Пути передачи – алиментарный, контактно-бытовой.Это мелкие палочковидные или кокковидные образования, окрашивающиеся по Романовскому—Гимзе в ярко-розовый цвет. Размножение микроорганизмов происходит в гистиоцитах и макрофагах, после разрушения которых отмечаются генерализация процесса и токсинемия. В процессе инфекции развивается реакция гиперчувствительности замедленного типа, формируется напряженный иммунитет. Заболевание характеризуется неясной клинической картиной. Диагностика:1) серологическое исследование (РСК, РПГА); 2) кожно-аллергическая проба (как ретроспективный метод диагностики). Специфическая профилактика: живая вакцина М-44.
V — группа пароксизмальных риккетсиозов, в которую входят:
1) волынская, или траншейная, лихорадка;
2) клещевой пароксизмальный риккетсиоз- Bartonella quintana.
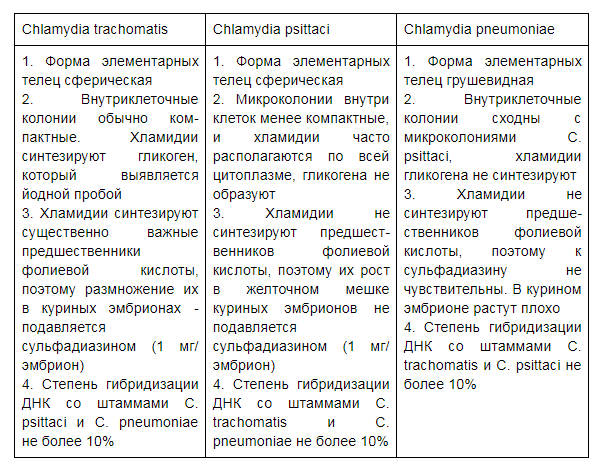
Хламидии (Chlamydia) -- мелкие грамотрицательные кокковидные бактерии. Все хламидии сгруппированы в порядок Chlamydiaceae, род Chlamydia, последний включает четыре вида: роды Chlamydia и Chlamydophila
-
Chlamydia trachomatis (Хламидия трахоматис).
-
Chlamydia psittaci.
-
Chlamydia pneumonia.
-
Chlamydia psittaci – вызывает у человека атипичную пневмонию, энцефаломиокардит, артрит, пиелонефрит.от птиц
-
Chlamydia pneumoniae вызывает у взрослых острые респираторные заболевания и мягкую форму атипичную пневмонии.
-
Вид Chlamydia trachomatis встречается только у человека, в ней выявлены 18 антигенных вариантов (серотипов). Серотипы А,В,С -- возбудители трахомы. Переносчиками являются насекомые, основной путь заражения -- попадание инфекционного агента посредством втирания в область слизистой оболочки глаза. Образующиеся в результате развития инфекционного процесса рубцы ведут к потере зрения. Серотипы L1-L3 размножаются в лимфоидной ткани и являются возбудителями тропической венерической болезни Lymphogranuloma venerum. В случае серотипов от D до К -- заражение происходит при половом контакте, значительно реже -- при втирании, новорожд енный при родах заражается от инфицированной матери.
Физиология и патогенез
Хламидии существуют в двух формах, различающихся по морфологическим и биологическим свойствам. Высокоинфекционной, спороподобной, внеклеточной формой является элементарное тельце (ЭТ), и вегетативной, репродуцирующейся, внутриклеточной -ретикулярное тельце (РТ). Первый этап инфекционного процесса -- адсорбция ЭТ на плазмалемме чувствительной клетки хозяина. путем эндоцитоза. Инвагинация участка плазмалеммы с адсорбированным ЭТ происходит в цитоплазму с образованием фагоцитарной вакуоли. Эта фаза занимает 7-10 часов. После этого уже в клетке в течение 6-8 часов происходит реорганизация ЭТ в вегетативную форму-ретикулярное тельце, способное к росту и делению. Именно на этой фазе эффективно курсовое применение антибактериальных препаратов. Размножение хламидий ведет к формированию включений, известных под названием телец Провачека. В течение 18-24 часов развития они локализованы в цитоплазматическом пузырьке, образованном из мембраны клетки хозяина. Далее начинается процесс созревания ретикулярных телец через переходные (промежуточные) тельца в течение 36-42 часа развития в ЭТ следующего поколения. Полный цикл репродукции хламидии равен 48-72 часам и завершается разрушением пораженной клетки, в случае возникновения для хламидии неблагоприятных метаболических условий этот процесс может затягиваться на более длительный период.
Хламидии могут высвобождаться из инфицированной клетки через узкий ободок цитоплазмы. При этом клетка может сохранять жизнеспособность, этим можно объяснить бессимптомность течения хламидийной инфекции. Хламидии поглощаются периферическими моноцитами и распространяются в организме, моноциты оседают в тканях и превращаются в тканевые макрофаги (в суставах, в сосудах, в области сердца). Тканевые макрофаги могут сохранять жизнеспособность в течение нескольких месяцев, являясь при этом мощным антигенным стимулятором, приводя к образованию фиброзных гранулем в здоровой ткани.
Микоплазмы. Таксономия. Характеристика. Микробиологическая диагностика. Лечение. Антропонозные бактериальные инфекции человека, поражающие органы дыхания или мочеполовой тракт. Микоплазмы относятся к классу Mollicutes, который включает 3 порядка: Acholeplasmatales, Mycoplasmatales, Anaeroplasmatales. Морфология: Отсутствие ригидной клеточной стенки, полиморфизм клеток, пластичность, осмотическую чувствительность, резистентность к различным агентам, подавляющим синтез клеточной стенки, в том числе к пенициллину и его производным. Грам «-», лучше окрашиваются по Романовскому—Гимзе; Биохимическая активность: Низкая. Выделяют 2 группы микоплазм: 1. разлагающие с образованием кислоты глюкозу, мальтозу, маннозу, фруктозу, крахмал и гликоген; 2.окисляющие глутамат и лактат, но не ферментирующие углеводы. Все виды не гидролизуют мочевину. Антигенная структура: Сложная, имеет видовые различия; основные АГ представлены фосфо- и гликолипидами, полисахаридами и белками; Факторы патогенности: адгезины, токсины, ферменты агрессии и продукты метаболизма. Адгезины входят в состав поверхностных АГ и обуславливают адгезию на клетках хозяина. Предполагают наличие нейротоксина у некоторых штаммов М. pneumoniae, так как часто инфекции дыхательных путей сопровождают поражения нервной системы. Эндотоксины выделены у многих патогенных микоплазм. У некоторых видов встречаются гемолизины. Эпидемиология: М. pneumoniaeколонизирует слизистую оболочку респираторного тракта; M. hominis, M. genitaliumuU. urealyticum— «урогенитальные микоплазмы» — обитают в урогенитальном тракте. Источник инфекции — больной человек. Механизм передачи — аэрогенный, основной путь передачи — воздушно-капельный. Внереспираторные проявления: гемолитическая анемия, неврологические расстройства, осложнения со стороны ССС.
Большинство видов — факультативные анаэробы; чрезвычайно требовательны к питательным средам и условиям культивирования. 12. Простейшие представлены 7 типами, из которых четыре типа (Sarcomastigophora, Apicomplexa, Ciliopkora, Microspora) включают возбудителей заболеваний у человека. Тип Sarcomastigophora. Подтип Mastigophora (жгутиконосцы) включает следующих патогенных представителей: трипаносому — возбудителя африканского трипаносомоза (сонная болезнь); лейшмании( L. tropica )— возбудителей кожной и висцеральной форм лейшманиозов; трихомонады, передающиеся половым путем и паразитирующие в толстой кишке человека; лямблию(Giardia intestinalis) — возбудителя лямблиоза. Эти простейшие характеризуются наличием жгутиков: один — у лейшмании, четыре свободных жгутика и короткая ундулирующая мембрана — у трихомонад. К подтипу Sarcodina (саркодовые) относится дизентерийная амеба — возбудитель амебной дизентерии человека. Морфологически сходна с ней непатогенная кишечная амеба. Эти простейшие передвигаются путем образования псевдоподий. Питательные вещества захватываются и погружаются в цитоплазму клеток. Половой путь размножения у амеб отсутствует. При неблагоприятных условиях они образуют цисту. 4
Тип Apicomplexa. В классе Sporozoa (споровики) патогенными представителями являются возбудители токсоплазмоза, кокцидиоза, саркоцистоза и малярии(Plasmodium falciparum). Жизненный цикл возбудителей малярии характеризуется чередованием полового размножения (в организме комаров Anopheles) и бесполого (в клетках тканей и эритроцитах человека они размножаются путем множественного деления). Токсоплазмы имеют форму полулуний. Токсоплазмозом человек заражается от животных. Токсоплазмы( Toxoplasma gondii,) в могут передаваться через плаценту и поражать центральную нервную систему и глаза плода. Тип Ciliophora. Патогенный представитель — возбудитель балантидиаза(Balantidium coli ) — поражает толстый кишечник человека. Балантидии имеют многочисленные реснички и поэтому подвижны. Тип Microspora включает микроспоридии — маленькие (0,5—10 мкм) облигатные внутриклеточные паразиты, широко распространенные среди животных и вызывающие у ослабленных людей диарею и гнойно-воспалительные заболевания.
13. Обмен веществ (метаболизм) разных живых организмов имеет сходные механизмы, но у микробов есть ряд особенностей:
-
Благодаря высокой интенсивности метаболизма вес перерабатываемых веществ в 30-40 раз больше веса самого микроорганизма.
-
В питании участвует вся поверхность клетки.
-
Пища перерабатывается выделяемыми ферментами снаружи, а внутрь клетки поступают образовавшиеся после этого более простые соединения.
Бактерии делятся на группы в зависимости от признака, по которому производится классификация:
По типам питания, в основе которых лежит источник азота и углерода, микроорганизмы подразделяются на две группы: автотрофы и гетеротрофы.
Автотрофы (от греческого autos – сам, tropha – питание) способны синтезировать сложные органические вещества, используя для этого простые неорганические соединения. Источником углерода и азота для этих микроорганизмов являются углекислота и другие неорганические соединения углерода, молекулярный азот воздуха и аммонийные соли. За счет этих простых соединений автотрофные микроорганизмы синтезируют белки, жиры, углеводы, витамины, ферменты.
Среди автотрофов (прототрофов) есть хемосинтезирующие, которые получают энергию за счет химических реакций, они лишены фотоситетических пигментов и фотосинтезирующие микроорганизмы, получающие энергию солнечного света и содержащие хлорофилоподобные пигменты.
К автотрофам относится меньшая часть микроорганизмов, большая же часть из них является гетеротрофами.
Гетеротрофы (от греческого heteros – другой, tropha – питание) способны ассимилировать углерод только из органических соединений. Что касается источника азота, то здесь могут быть разные источники. Многие гетеротрофы усваивают азот из аммонийных соединений. Есть представители гетеротрофных микробов, которые в качестве источника азота используют аминокислоты, а некоторые (преимущественно патогенные виды) используют нативный (неизмененный) белок.
-
По используемому источнику энергии:
-
фототрофы – энергия солнечного света; а)хлорофилл- с выделением кислорода (цианобактерий )
б)бактериохлорофилл- кислорода не происходит
-
хемотрофы – энергия окислительно-восстановительных реакций- большенство болезнеторных бактерий (Химические реакции могут быть двух видов: аэробными, с обязательным присутствием кислорода или анаэробными, то есть бескислородными. Процессы первого типа принято называть дыханием, а второго – брожением.)
По типу соединения, служащего донором электронов:
-
органотрофы – органические вещества- Pseudomonas fluorescens
-
литотрофы – неорганические вещества.
По источнику углерода:
-
автотрофы – углекислый газ;неорганика
-
гетеротрофы – органические вещества. паразитов и сапрофитов
Хемоорганоавтотрофы. Окисляют трудноусваиваемые вещества. Например, некоторые представители аминобактерий (Aminobacter), метилобактерий (Methylobacterium), флавобактерий (Flavobacterium), псевдомонад (Pseudomonas).
Хемоорганогетеротрофы. Большинство видов бактерий
Условно можно выделить четыре механизма проникновения питательных веществ в бактериальную клетку: простая диффузия, облегченная диффузия, активный транспорт, транслокация групп. Наиболее простой механизм поступления веществ в клетку простая диффузия, при которой перемещение веществ происходит вследствие разницы их концентрации по обе стороны цитоплазматической мембраны. без затраты энергии. Облегченная диффузия происходит также в результате разницы концентрации веществ по обе стороны цитоплазматической мембраны. Однако этот процесс осуществляется с помощью молекул-переносчиков, локализующихся в цитоплазматической мембране и обладающих специфичностью.. Облегченная диффузия протекает без затраты энергии, вещества перемещаются из области с более высокой концентрацией в область с более низкой. Активный транспорт происходит с помощью пермеаз и направлен на перенос веществ меньшей концентрации в сторону большей, то есть как бы против течения, поэтому данный процесс сопровождается затратой метаболической энергии (АТФ), образующейся в результате окислительновосстановительных реакций в клетке. Перенос (транслокация) групп сходен с активным транспортом, но отличается тем, что переносимая молекула видоизменяется в процессе переноса, например фосфорилируется. Выход веществ из клетки осуществляется за счет диффузии и при участии транспортных систем. 14 Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с образованием АТФ-универсального аккумулятора химической энергии. Энергия необходима микробной клетке для ее жизнедеятельности. При дыхании происходят процессы окисления и восстановления: окисление — отдача донорами (молекулами или атомами) во-дорода или электронов; восстановление — присоединение водорода или электронов к акцептору. Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат (такое дыхание называется анаэробным — нитратным, сульфатным, фумаратным). Анаэробиоз (от греч. аег — воздух + bios — жизнь) — жизнедеятельность, протекающая при отсутствии свободного кислорода. Если донорами и акцепторами водорода являются органические соединения, то такой процесс называется брожением. При брожении происходит ферментативное расщепление органических соединений, преимущественно углеводов, в анаэробных условиях. С учетом конечного продукта расщепления углеводов различают спиртовое, молочнокислое, уксуснокислое и другие виды брожения. По отношению к молекулярному кислороду бактерии можно разделить на три основные группы: облигатные, т.е. обязательные, аэробы, облигатные анаэробы и факультативные анаэробы. Методы культивирования анаэробов. Для культивирования анаэробов необходимо понизить окислительно-восстановительный потенциал среды, создать условия анаэробиоза, т. е. пониженного содержания кислорода в среде и окружающем ее пространстве. Это достигается применением физических, химических и биологических методов. Физические методы. Основаны на выращивании микроорганизмов в безвоздушной среде, что достигается: 1) посевом в среды, содержащие редуцирующие и легко окисляемые вещества; 2) посевом микроорганизмов в глубину плотных питательных сред; 3) механическим удалением воздуха из сосудов, в которых выращиваются анаэробные микроорганизмы; 4) заменой воздуха в сосудах каким-либо индифферентным газом. В качестве редуцирующих веществ обычно используют кусочки (около 0,5 г) животных или растительных тканей (печень, мозг, почки, селезенка, кровь, картофель, вата). Эти ткани связывают растворенный в среде кислород и адсорбируют бактерии. Чтобы уменьшить содержание кислорода в питательной среде, ее перед посевом кипятят 10—15 мин, а затем быстро охлаждают и заливают сверху небольшим количеством стерильного вазелинового масла. Высота слоя масла в пробирке около 1 см. В качестве легко окисляемых веществ используют глюкозу, лактозу и муравьинокислый натрий. Лучшей жидкой питательной средой с редуцирующими веществами является среда Китта — Тароцци, для накопления анаэробов при первичном посеве из исследуемого материала и для поддержания роста выделенной чистой культуры анаэробов.
Посев микроорганизмов в глубину плотных сред производят по способу который состоит в механической защите посевов анаэробов от кислорода воздуха. Берут стеклянную трубку длиной 30 см и диаметром 3—6 мм. Один конец трубки вытягивают в капилляр в виде пастеровской пипетки, а у другого конца делают перетяжку. В оставшийся широкий конец трубки вставляют ватную пробку. В пробирки с расплавленным и охлажденным до 50°С питательным агаром засевают исследуемый материал. Затем насасывают засеянный агар в стерильные трубки Виньяль — Вейона. Капиллярный конец трубки запаивают в пламени горелки и трубки помещают в термостат. Так создаются благоприятные условия для роста самых строгих анаэробов. Для выделения отдельной колонии трубку надрезают напильником, соблюдая правила асептики, на уровне колонии,а колонию захватывают стерильной петлей и переносят в пробирку с питательной средой для дальнейшего выращивания и изучения в чистом виде. Удаление воздуха производят путем его механического откачивания из специальных приборов — анаэростатов, в которые помещают чашки с посевом анаэробов. Переносный анаэростат представляет собой толстостенный металлический цилиндр с хорошо притертой крышкой (с резиновой прокладкой), снабженный отводящим краном и вакуумметром. После размещения засеянных чашек или пробирок воздух из анаэростата удаляют с помощью вакуумного насоса. Замену воздуха индифферентным газом (азотом, водородом, аргоном, углекислым газом) можно производить в тех же анаэростатах путем вытеснения его газом из баллона. Химические методы. Основаны на поглощении кислорода воздуха в герметически закрытом сосуде (анаэростате, эксикаторе) такими веществами, как пирогаллол или гидросульфит натрия. Биологические методы. Основаны на совместном выращивании анаэробов со строгими аэробами. Для этого из застывшей агаровой пластинки по диаметру чашки вырезают стерильным скальпелем полоску агара шириной около 1 см. Получается два агаровых полудиска в одной чашке. На одну сторону агаровой пластинки засевают аэроб, например часто используют S. aureus. На другую сторону засевают анаэроб. Края чашки заклеивают пластилином или заливают расплавленным парафином и помещают в термостат. При наличии подходящих условий в чашке начнут размножаться аэробы. После того, как весь кислород в пространстве чашки будет ими использован, начнется рост анаэробов (через 3—4 сут). В целях сокращения воздушного пространства в чашке питательную среду наливают возможно более толстым слоем. Комбинированные методы. Основаны на сочетании физических, химических и биологических методов создания анаэробиоза.
15. Рост и размножение бактерий- Жизнедеятельность бактерий характеризуется ростом — формированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размножением — самовоспроизведением, приводящим к увеличению ко-личества бактериальных клеток в популяции. Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут раз-множаться спорами.,путем фрагментации нитевидных клеток. Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а грамотрицательные — путем перетяжки, в результате образования гантелевид-ных фигур, из которых образуются две одинаковые клетки. Делению клеток предшествует репликация бактериальной хромосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной нитью), приводящая к удвоению молекул ДНК бактериального ядра — нуклеоида. Репликация ДНК происходит в три этапа: инициация, элонгация, или рост цепи, и терминация Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не изменяющийся объем питательной среды, размножаясь, потребляют питательные элементы, что приводит в дальнейшем к истощению питательной среды и прекращению роста бактерий. Культивирование бактерий в такой системе называют периодическим культивированием. Если же условия культивирования поддерживаются путем непрерывной подачи свежей питательной среды и оттока такого же объема культуральной жидкости, то такое культивирование называется непрерывным, При выращивании бактерий на жидкой питательной среде наблюдается придонный(стрептококи) диффузный( Yersinia pestis- пленка с тяжами) или поверхностный (в виде пленки(бациллы, микобактерии и вибрионы ) рост культуры. Рост периодической культуры бактерий, выращиваемых на жидкой питательной среде, подразделяют на несколько фаз, или периодов: 1. лаг-фаза; 2. фаза логарифмического роста; 3. фаза стационарного роста, или максимальной концентрации бактерий; 4. фаза гибели бактерий.
Размножение
бактерий на плотной питательной среде.
Бактерии, растущие на плотных
питательных средах, образуют изолированные
колонии округлой формы с ровными или
неровными краями (S- и R-формы), различной
консистенции и цвета, зависящего от
пигмента бактерий. Пигменты, растворимые
в воде, диффундируют в питательную среду
и окрашивают еѐ. Другая группа пигментов
нерастворима в воде, но растворима в
органических растворителях. И, наконец,
существуют пигменты, не растворимые ни
в воде, ни в органических соединениях.
Наиболее распространены среди
микроорганизмов такие пигменты, как
каротины, ксантофиллы и меланины.
Меланины являются нерастворимыми
пигментами черного, коричневого или
красного цвета, синтезирующимися из
фенольных соединений. Меланины наряду
с каталазой, супероксидцисмутазой и
пероксидазами защищают микроорганизмы
от воздействия токсичных перекисных
радикалов кислорода. Если
плотность засева большая, то бактерии
формируют на поверхности агара сплошной
налет – так называемый, «рост газоном»
или «сливной рост».
2.
Если засев проводится таким образом,
что каждая бактериальная клетка лежит
на поверхности агара на большом расстоянии
от других, то, после многократных делений
она формирует изолированную колонию
(говорят еще об «изолированном росте»).
А так, как колония – результат размножения
од-ной клетки, то ее, с некоторыми
допущениями, рассматривают как клональную
культуру. Именно из материала отдельной,
изолированной, колонии в процессе
культурального метода исследования
полу-чают так называемую «чистую
культуру» – культуру, содержащие клетки
только одного вида.

16.
В основе всех метаболических реакций
в бактериальной клетке лежит деятельность
ферментов, которые принадлежат к 6
клас-сам: оксиредуктазы, трансферазы,
гидролазы, лигазы, лиазы, изомеразы.
Ферменты, образуемые бактериальной
клеткой, могут локализоваться как внутри
клетки — эндоферменты.
пищеварительные ферменты, которые
расщепляют сложные питательные вещества
до простых веществ; б) защитные ферменты,
например, пенициллиназа защищает
клеточную стенку от действия антибиотика
пенициллина; в) ферменты
агрессии –
факторы вирулентности патогенных
бактерий; гиалуронидаза –
расщепляет гиалуроновую
кислоту;- дезоксирибонуклеаза –
расщепляет ДНК клеток;- фибринолизин –
расщепляет коллаген;- плазмокоагулаза –
свертывает плазму крови; нейраминидаза –
расщепляет нейраминовую кислоту;)это
экзоферменты. Большинство гидролаз
является экзоферментами, которые,
выделяясь в окружающую среду, расщепляют
крупные молекулы пептидов, полисахаридов,
липидов до мономеров и димеров, способных
проникнуть внутрь клетки. Ряд экзоферментов,
например гиалуронидаза, коллагеназа и
другие, являются ферментами агрессии.
Некоторые ферменты локализованы в
периплазматическом пространстве
бактериальной клетки. Они участвуют в
процессах переноса веществ в бактериальную
клетку.
Идентификация бактерий по
ферментативной активности. Наиболее
часто определяют ферменты класса
гидролаз и оксидоредуктаз, используя
специальные методы и среды. Для определения
протеолитической
активности микроорганизмы засевают в
столбик желатина уколом. Через 3—5 дней
посевы просматривают и отмечают характер
разжижения желатина. При разложении
белка некоторыми бактериями могут
выделяться специфические продукты —
индол, сероводород, аммиак. Для их
определения служат специальные
индикаторные бумажки, ко-торые помещают
между горлышком и ватной пробкой в
пробирку с МПБ или (и) пептонной водой,
засеянными изучаемыми микроорганизмами.
Индол (продукт разложения триптофана)
окрашивает в розовый цвет полоску бумаги
с раствором щавелевой кислоты. Бумага,
пропитанная раствором ацетата свинца,
в присутствии сероводорода чернеет.
Для определения аммиака используют
красную лакмусовую бумажку. Для многих
микроорганизмов таксономическим
признаком служит способность разлагать
определенные углеводы с образованием
кислот и газообразных продуктов. Для
выявления этого используют среды Гисса,
содержащие различные углеводы (глюкозу,
сахарозу, мальтозу, лактозу и маннит).
Для обнаружения кислот в среду добавлен
реактив Андреде, который изменяет свой
цвет от бледно-желтого до красного в
интервале рН 7,2—6,5, поэтому набор сред
Гисса с ростом микроорганизмов называют
«пестрым рядом». Гидролиз мочевины
определяют по выделению аммиака
(лакмусовая бумажка) и подщелачиванию
среды. Для определения нитритов
используют реактив Грисса: Появление
красного
окрашивания свидетельствует о наличии
нитритов.


17. Выведенные из питательной среды новые виды бактерий, свойства которых ещё не определены, называются чистой культурой. После окончательной идентификации их характеристик, бактерии, выведенные из определённого места и в определённое время, получают название штамм. При этом допускается незначительное различие в свойствах, месте или времени выделения штамма одного вида. Чистой культурой называется популяция бактерий одного вида или одной разновидности, выращенная на питательной среде. Многие виды бактерий подразделяют по одному признаку на биологические варианты — биовары. Биовары, различающиеся по биохимическим свойствам, называют хемоварами, по антигенным свойствам — сероварами, по чувствительности к фагу — фаговарами. Методы выделения чистых культур бактерий. Универсальным инструментом для производства посевов является бактериальная петля. Кроме нее, для посева уколом при-меняют специальную бактериальную иглу, а для посевов на чашках Петри — металлические или стеклянные шпатели. Для посевов жидких материалов наряду с петлей используют пастеровские и градуированные пипетки. Первые предварительно из-готовляют из стерильных легкоплавких стеклянных трубочек, которые вытягивают на пламени в виде капилляров. Конец ка-пилляра сразу же запаивают для сохранения стерильности. У пастеровских и градуированных пипеток широкий конец за-крывают ватой, после чего их помещают в специальные пеналы или обертывают бумагой и стерилизуют. 19-20. Бактериофаги — вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродуцироваться в них и вызывать их растворение (лизис). Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги. Вирулентные фаги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Про-цесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодей-ствия вирусов человека и животных с клеткой хозяина. Однако для фагов, имеющих хвостовой отросток с сокращающимся чехлом, он имеет особенности. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе «прокалывания» клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и активно впрыскивается в цитоплазму клетки.. После биосинтеза фаговых компонентов и их самосборки Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий. Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков. Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бакте-рий, содержащая профаг, получила название лизогенной. Практическое применение фагов. Бактериофаги используют в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т. е. определении фаговара (фаготипа). Для этого применяют метод фаготипирования, основанный на строгой специфичности действия фагов: на чашку с плотной питательной средой, засеянной «газоном» чистой культурой возбудителя, наносят капли различных диагностических типоспецифических фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее лизис (образование стерильного пятна, «бляшки», или «негативной колонии», фага). Методику фаготипирования используют для выявления источника и путей распространения инфекции (эпидемиологическое маркирование). Выделение бактерий одного фаговара от разных больных указывает на общий источник их заражения. По содержанию бактериофагов в объектах окружающей среды (например, в воде) можно судить о присутствии в них соответствующих патогенных бактерий. Подобные исследования проводят при эпидемиологическом анализе вспышек инфекционных болезней. Фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги и комбинированные препараты (колипротейный, пиобактериофаги и др). Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблети-рованных форм, свечей или аэрозолей. Бактериофаги широко применяют в генной инженерии и биотехнологии в качестве векторов для получения рекомбинантных ДНК. Выделение фага из объектов окружающей среды
Для получения вирулентного фага готовят фильтрат, пропуская исходный материал (вода, суспензия фекалий и др.) через бактериальные фильтры. Фильтрат вместе с соответствующей бактериальной культурой засевают в бульон и инкубируют при 37 °С в течение 18 - 24 ч. После лизиса культуры оставшиеся бактериальные клетки удаляют центрифугированием или фильтрацией через бактериальный фильтр. Наличие фага в фильтрате определяют качественными и количественными методами.[3]
Качественный метод определения фагов E.coli
Чашку Петри с питательным агаром засевают суточной бульонной культурой кишечной палочки газоном и подсушивают при 37 °С в течение 10—15 мин. Затем на поверхность газона наносят каплю фага и наклоняют так, чтобы капля стекла к противоположному краю. После суточной инкубации в термостате просматривают чашку, отмечая наличие зоны лизиса по месту стекания капли фага.[3]
Количественный метод - определение титра фага по методу Грациа
готовят смесь из следующего разведения фага (10-6) с бактериями и полужидким агаром и выливают на поверхность агара в другой чашке, затем - из разведения 10-5. После застывания второго слоя агара чашки инкубируют при 370 С, затем подсчитывают число негативных колоний фага. Число этих колоний соответствует количеству фаговых частиц в засеянной смеси. Исходя из него, можно вычислить количество пятнообразующих единиц в 1 мл исходной суспензии фага. Эта величина, характеризующая концентрацию фага, называется его титром Фенотипические проявления фаговой конверсии очень разнообразны. Ею обусловлена способность к синтезу токсинов у Clostridium botulinum, Clostridium tetani, Corynebacterium diphtheriae и Corynebacterium ulcerans, изменение фаготипов и продукция гемолизинов у Staphylococcus aureus, синтез новых компонентов антигенов у различных групп Salmonella.
22.
генотип.фенотип. наследственность



23. Изменчивость микроорганизмов подразделяется на наследственную, обусловленную генотипическими изменениями, и ненаследственную (фенотипическую). Фенотипические изменения При фенотипической изменчивости микробы, образовавшиеся из одной материнской клетки, могут различаться между собой по ферментативной активности, морфологическим признакам, потребности в источниках питания. К фенотипической изменчивости относятся: Адаптация – приспособление микроорганизмов к новым условиям среды. Диссоциация – культурная изменчивость, когда, например, из засеянной на плотную среду чистой культуры вырастают резко отличающиеся по морфологической структуре колонии (тип S – гладкие, тип R – шероховатые, тип M – слизистые). Модификация – изменение микроорганизмов под влиянием условий среды. Изменяются только фенотипические (внешние) признаки (форма, размеры, цвет колоний). Модификация наблюдается в нормальных условиях жизни, это реакция на внешние раздражения, не связанные с нарушением физиологических процессов в организме. Модификационные изменения легко исчезают при устранении условий, их вызвавших.
Наследственная (генотипическая) изменчивость, связанная с мутациями, — мутационная изменчивость. Основу мутации со-ставляют изменения последовательности нуклеотидов в ДНК, полная или частичная их утрата, т. е. происходит структурная перестройка генов, проявляющаяся фенотипически в виде измененного признака. Наследственная изменчивость, связанная с рекомбинациями, называется рекомбинационной изменчивостью. Мутации – внезапные, скачкообразные изменения генов. Процесс мутирования генов приводит к таким изменениям, которые передаются по наследству и сохраняются даже тогда, когда вызвавший их фактор перестает действовать.
24. мутации