

3.3.8. Синтез днк
Различными методами показано, что в период, предшествующий делению клеток, количество ДНК в них удваивается. При этом синтез ДНК осуществляется таким образом, что и в дочерних, и в материнских клетках содержатся совершенно идентичные молекулы ДНК. Это означает, что в процессе синтеза реализуется механизм воспроизведения структуры ДНК, который обеспечивает передачу наследственных свойств в ходе размножения клеток. Механизм образования в клетках организмов новых молекул ДНК, которые идентичны по генетическим свойствам уже имеющимся молекулам, называют репликацией ДНК.
О
–А–Т–Г–Ц–Т–Г–А–Ц–
–Т–А–Ц–Г–А–Ц–Т–Г–
– А
– Т – Г – Ц – Т – Г – А – Ц –
–А–Т–Г–Ц–Т–Г–А–Ц–
‥ ‥ …… ‥ … ‥ …
‥ ‥ …… ‥ … ‥ …
– Т
– А – Ц – Г – А – Ц – Т – Г –
–Т–А–Ц–Г–А–Ц–Т–Г–
–А–Т–Г–Ц–Т–Г–А–Ц–
комплементарные
–Т–А–Ц–Г–А–Ц–Т–Г– синтез
новых
цепи исходной
разъединенные комплементарных
молекулы ДНК
цепи ДНК цепей ДНК
Синтез ДНК катализируют ферменты ДНК-нуклеотидилтрансфе-разы, или более кратко ДНК-полимеразы. Эти ферментные белки содержат в своём составе семь разных субъединиц, образующих олигомерную молекулу с молекулярной массой около 500 тыс. ДНК-полимеразы осуществляют синтез ДНК на одноцепочечной матрице, если на ней уже содержится небольшой олигонуклеотидный фрагмент, комплементарный нуклеотидным остаткам матрицы и называемый затравкой, у которой на 3'-конце имеется свободная гидроксильная группа. Субстратами для построения синтезируемой полинуклеотидной цепи ДНК служат дезоксирибонуклеозидтрифосфаты, т.е. дАТФ, дГТФ, дЦТФ, дТТФ. Порядок присоединения фосфорилированных нуклеотидных остатков определяется их комплементарным взаимодействием с соответствующими нуклеотидными остатками на ДНК-матрице, с которыми они образуют водородные связи. На активирование ДНК-полимеразы затрачивается энергия, которая высвобождается при гидролизе макроэргической связи АТФ с образованием АДФ и фосфорной кислоты.
В ходе полимеразной реакции осуществляется
гидролиз макроэргической связи
дезоксинуклеозидтрифосфата и высвобождение
пирофосфата, а нуклеотидный остаток в
виде дезоксинуклеотидмонофосфата
присоединяется к 3'-ОН затравки. Так
последовательно происходит наращивание
синтезируемой полинуклеотидной цепи
ДНК в направлении от 5'-конца к 3'-концу.
Вновь синтезируемая цепь ДНК комплементарна
и антипаралельна полинуклеотидной цепи
матрицы, что отвечает принципам
постро-ения двойной спирали ДНК.
Последовательность присоединения
нуклеотидных остатков в ходе матричного
синтеза ДНК можно представить следующим
образом:
ходе полимеразной реакции осуществляется
гидролиз макроэргической связи
дезоксинуклеозидтрифосфата и высвобождение
пирофосфата, а нуклеотидный остаток в
виде дезоксинуклеотидмонофосфата
присоединяется к 3'-ОН затравки. Так
последовательно происходит наращивание
синтезируемой полинуклеотидной цепи
ДНК в направлении от 5'-конца к 3'-концу.
Вновь синтезируемая цепь ДНК комплементарна
и антипаралельна полинуклеотидной цепи
матрицы, что отвечает принципам
постро-ения двойной спирали ДНК.
Последовательность присоединения
нуклеотидных остатков в ходе матричного
синтеза ДНК можно представить следующим
образом:
ДНК-полимеразы осуществляют синтез полинуклеотидных цепей ДНК с очень высокой скоростью. Например, в клетках бактерий скорость этого процесса составляет около 1000 нуклеотидных остатков за 1 с, а в клетках высших организмов – несколько десятков нуклеотидных остатков (около 100). Однако даже при такой высокой скорости синтеза фермент действует очень точно и вероятность присоединения неправильного нуклеотидного остатка обычно не превышает 10-6–10-8, т.е. одна ошибка на десятки миллионов нуклеотидных остатков.
ДНК-полимераза с высокой точностью присоединяет к 3'-концу растущей цепи ДНК нуклеотидные остатки, которые комплементарны нуклеотидам матрицы. При этом правильность присоединения каждого нуклеотидного остатка контролируется ферментом дважды. Первый раз, когда присоединяется очередной нуклеотидный остаток, и второй раз, когда присоединяется следующий нуклеотидный остаток. Если предшествующий нуклеотидный остаток не комплементарен матрице, то следующая фосфоэфирная связь не образуется до тех пор, пока неправильный нуклеотидный остаток не будет удалён и его место не займёт правильная нуклеотидная группировка. В связи с этим некоторые ДНК-полимеразы не только способны катализировать синтез ДНК, но и обладают 3'-экзонуклеазной активностью, которая обеспечивает удаление некомплементарного матрице нуклеотидного остатка на 3'-конце синтезируемой цепи ДНК.
Как было указано ранее, ДНК-полимеразы присоединяют нуклеотидные остатки в ходе синтеза ДНК к 3'-концам олигонуклеотида, называемого «затравкой», комплементарно присоединённого к матрице. При этом образование «затравки» катализирует специальный фермент, который назвали ДНК-праймазой. Этот фермент катализирует присоединение к мат-ричной цепи ДНК нескольких остатков рибонуклеотидов, формирующих короткую затравочную цепь РНК, которая комплементарна матричной цепи ДНК и образует с ней двойную спираль. Именно к 3'-концу РНК-за-травки присоединяются далее остатки дезоксирибонуклеотидов с помощью ДНК-полимеразы. После того как синтез новой ДНК-цепи заканчивается, РНК-затравки удаляются специальной ДНК-полимеразой, которая проявляет 5'-экзонуклеазную активность, т.е. отщепляет от полинуклеотидной цепи 5'-концевые рибонуклеотидные остатки.
У бактерий репликация ДНК начинается в определённом участке хромосомы, который сокращённо обозначают ori (от английского слова origin – начало). Этот участок хромосомы имеет специфическую последовательность нуклеотидных остатков, узнаваемую одним из белков, участвующим в инициации синтеза ДНК. Его называют инициаторным белком. В хромосомах высших организмов репликация начинается одновременно на многих участках, расстояние между которыми составляет порядка 100 тыс. нуклеотидных остатков.
П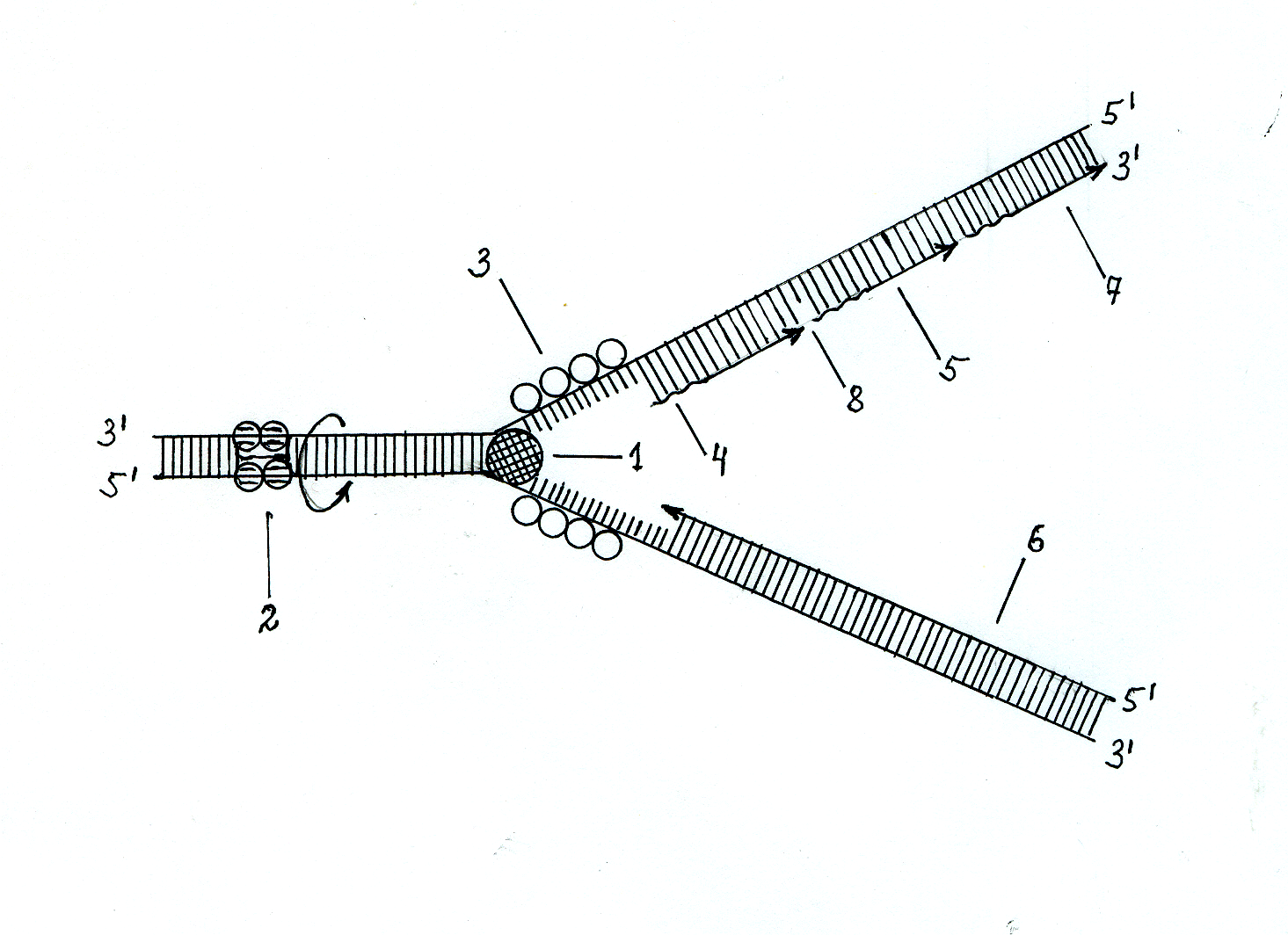 осле
того как инициаторный белок связывается
с участком ДНК ori, к
этому участку присоединяется в
определённой последовательности
комплекс из шести белков, главным из
которых является хеликаза. Это –
АТФ-зависимый фермент, использующий
энергию гидролиза АТФ для разъединения
полинуклеотидных цепей двойной спирали
ДНК. Ферментный белок хеликазы,
присоединившись к одной из цепей ДНК,
движется вдоль этой цепи и расплетает
перед собой двойную спираль ДНК, образуя
репликативную вилку, которая включает
участок двойной спирали ДНК и две
отходящие от него одноцепочечные ветви
(рис. 3.11).
осле
того как инициаторный белок связывается
с участком ДНК ori, к
этому участку присоединяется в
определённой последовательности
комплекс из шести белков, главным из
которых является хеликаза. Это –
АТФ-зависимый фермент, использующий
энергию гидролиза АТФ для разъединения
полинуклеотидных цепей двойной спирали
ДНК. Ферментный белок хеликазы,
присоединившись к одной из цепей ДНК,
движется вдоль этой цепи и расплетает
перед собой двойную спираль ДНК, образуя
репликативную вилку, которая включает
участок двойной спирали ДНК и две
отходящие от него одноцепочечные ветви
(рис. 3.11).
Рис. 3.11. Синтез ДНК в репликативной вилке
1 – хеликаза; 2 – топоизомераза; 3 – SSB-белки; 4 – РНК-затравки; 5 – фрагменты Оказаки; 6 – ведушая цепь синтезируемой ДНК; 7 - отстающая цепь синтезируемой ДНК; 8 – место действия ДНК-лигазы.
Самопроизвольное объединение в двойную спираль образовавшихся под действием хеликазы одноцепочечных участков ДНК предотвращают SSB-белки (от английского single strand binding proteins), которые имеют избирательное сродство к разъединённым полинуклеотидным цепям ДНК. Эти белки, вплотную прилегая друг к другу, связывают одноцепочечные участки ДНК и делают их более реакционноспособными для взаимодействия с ферментом ДНК-полимеразой.
В результате расплетания хеликазой двойной спирали ДНК возникает её вращательное движение впереди репликативной вилки. Однако в связи с тем, что хромосома связана и закреплена клеточными мембранами, вращение двуспиральной ДНК приводит к образованию дополнительных витков и сверхскрученных узлов на пути продвижения репликативной вилки, что могло бы в дальнейшем остановить процесс репликации ДНК. Но этого не происходит вследствие того, что на участке сверхспирализации впереди по движению репликативной вилки действует фермент топоизомераза, образующий разрыв фосфоэфирной связи в одной из цепей двуспиральной ДНК. За счёт такого действия топоизомеразы обеспечивается свободное вращение оставшейся неразорванной одноцепочечной нити ДНК вокруг её σ-связи, что приводит к снятию сверхспирализации ДНК на участке продвижения репликативной вилки и раскручиванию образовавшихся узлов, вызванных действием фермента хеликазы. После раскручивания дополнительных витков сверхспирализации топоизомера восстанавливает разорванную фосфоэфирную связь и таким образом сохраняет структурную целостность двойной спирали ДНК, которая участвует в процессе репликации.
В результате действия хеликазы и SSB-белков образуются две разъединённые цепи ДНК, имеющие противоположную направленность. Одна из них начинается от 3'-конца, другая – от 5'-конца. На цепи ДНК, начинающейся с 3'-конца, после образования РНК-затравки возможен непрерывный синтез новой цепи ДНК под действием фермента ДНК-полиме-разы, способного присоединять к 3'-ОН растущей цепи новые нуклеотидные остатки. Однако на цепи ДНК, начинающейся с 5'-конца, такой синтез новой цепи ДНК невозможен. Он должен происходить в обратном направлении после образования одноцепочечного участка ДНК определённой длины, вследствие чего синтез ДНК на одноцепочечной ветви репликативной вилки, начинающейся с 5'-конца, происходит медленнее, чем на другой одноцепочечной ветви, начинающейся с 3'-конца. В связи с этим одну из синтезирующихся цепей ДНК называют ведущей, а другую цепь, синтез которой происходит медленнее, – отстающей.
В опытах Р. Оказаки было показано, что отстающая цепь ДНК синтезируется в виде коротких фрагментов, называемых фрагментами Оказаки. Каждый такой фрагмент начинается с синтеза РНК-затравки с участием ДНК-праймазы, а далее действует фермент ДНК-полимераза, который уже синтезирует ДНК-цепь фрагмента Оказаки, доводя её до следующей РНК-затравки. После этого в действие включается фермент ДНК- полимераза, обладающая 5'-экзонуклазной активностью, которая удаляет впереди себя РНК-затравку и вместо неё достраивает цепь ДНК до следующего фрагмента Оказаки. На стыке двух фрагментов Оказаки имеется с одной стороны 3'-ОН, а с другой 5'-фосфатная группа, между которыми под действием фермента ДНК-лигазы образуется фосфодиэфирная связь, соединяющая фрагменты Оказаки в единую синтезируемую цепь ДНК.
Линейные размеры фрагментов Оказаки зависят от вида организмов; у бактерий они составляют в длину до 1000 нуклеотидных остатков, у высших организмов – около 100.
В основных чертах механизм репликации ДНК у разных организмов имеет много общего, хотя у высших организмов по сравнению с низшими имеются определённые особенности. Одна из таких особенностей связана со строением хромосом. У высших организмов хромосомы линейны и у них на 5'-конце отстающей цепи ДНК остаётся недореплицированный участок в связи с неспособностью ДНК-полимераз инициировать синтез ДНК на одноцепочечной матрице. Для того чтобы репликация ДНК проходила полностью, у высших организмов имеются удлинения на концах хромосом из повторяющихся последовательностей нуклеотидных остатков, которые образуют изгиб с разворотом цепи на 180º, так что свободных концов у хромосомы фактически нет. Присоединение к концам хромосом повторяющихся последовательностей нуклеотидных остатков катализирует специальная трансфераза. Длина концевых фрагментов хромосом контролируется регуляторной системой клетки.
Необходимо отметить, что репликация ДНК – энергоёмкий процесс, который во время своего осуществления вызывает активизацию биоэнергетических превращений, сопряжённых с синтезом АТФ и других макроэргических соединений. Энергия АТФ затрачивается на новообразование дезоксирибонуклеозидфосфатов, участвующих в синтезе полинуклеотидных цепей ДНК; рибонуклеозидтрифосфатов, участвующих в синтезе РНК-затравок; на работу хеликаз и топоизомераз, а также активацию ДНК-полимераз, которая происходит при переходе этого фермента на каждый из фрагментов Оказаки при синтезе отстающей цепи ДНК.
Как было указано ранее, репликация ДНК в клетках организмов осуществляется с очень высокой точностью, однако с небольшой частотой ошибки всё-таки происходят, и в структуру новосинтезируемой цепи ДНК включаются нуклеотидные остатки, которые не комплементарны матрице. Такие ошибки репликации исправляют ферменты, относящиеся к системе репарации ДНК. Процесс репарации ДНК – это исправление ошибок репликации ДНК и устранение изменённых нуклеотидных остатков, которые возникают под воздействием мутагенных факторов – радиации, ультрафиолетового излучения, алкилирующих химических веществ, дезаминирования и потери оснований в нуклеотидных остатках.
В систему репарации ДНК входят ферменты, способные расщеплять или восстанавливать межнуклеотидные фосфодиэфирные связи, удалять изменённые неправильно присоединённые нуклеотидные остатки и присоединять вместо них правильные нуклеотидные остатки, которые комплементарны матричной цепи ДНК. На репарационные процессы затрачивается значительное количество энергии, которая обеспечивается за счёт гидролиза АТФ.
Функционирование системы репарации ДНК обеспечивает исключение из её структуры неправильно присоединённых или изменённых под действием мутагенных факторов нуклеотидных остатков и таким образом устойчиво поддерживает стабильность содержащейся в молекулах ДНК генетической информации. Однако с небольшой частотой иногда такие ошибки возникают и дают начало при репликации изменённым молекулам ДНК. Появление в результате ошибок репликации или действия мутагенных факторов молекул ДНК с изменённым составом нуклеотидных остатков получило название генетических мутаций.