

3.1. Содержание и соотношение азотистых оснований в днк различных организмов
|
Виды организмов |
Содержание оснований, моль % |
Отношение А + Т/Г + Ц |
|||
|
А |
Т |
Г |
Ц |
||
|
Человек |
30,9 |
29,4 |
19,9 |
19,8 |
1,52 |
|
Овца |
29,3 |
28,3 |
21,4 |
21,0 |
1,36 |
|
Черепаха |
29,7 |
27,9 |
22,0 |
21,3 |
1,31 |
|
Зародыши пшеницы |
27,3 |
27,1 |
22,7 |
22,8 |
1,19 |
|
Листья моркови |
26,7 |
26,8 |
23,2 |
23,3 |
1,15 |
|
Дрожжи |
31,3 |
32,9 |
18,7 |
17,1 |
1,79 |
|
Бактерии: Еscherichia coli Clostridium perfringens |
24,7 36,9 |
23,6 36,3 |
26,0 14,0 |
25,7 12,8 |
0,93 2,70 |
|
Бактериофаг Т 7 |
26,0 |
26,0 |
24.0 |
24,0 |
1.08 |
Изучение нуклеотидного состава ДНК различных организмов (табл. 3.1) показало, что соотношение пар азотистых оснований А + Т/Г + Ц является важным показателем специфичности ДНК у разных организмов. Каждый вид организмов имеет свойственный ему нуклеотидный состав ДНК. Нуклеотидный состав ДНК у одного и того же генотипа остаётся постоянным в изменяющихся условиях окружающей среды, что является важнейшим фактором сохранения наследственных свойств организма.
В дальнейшем было установлено, что специфичность молекул ДНК, свойственная конкретному генотипу, определяется последовательностью соединения остатков нуклеотидов в полинуклеотидной цепи, которую называют первичной структурой ДНК.
Последовательность нуклеотидных остатков в полинуклеотидной це-пи записывают в виде сокращённых обозначений нуклеотидов слева направо, начиная от 5'-конца. Буквенные обозначения нуклеотидов могут быть соединены чёрточками. Например, в нуклеотидной последовательности Т–Ц–Г–А–Ц–А слева направо последовательно соединены фосфодиэфирными связями остатки дТМФ, дЦМФ, дГМФ, дАМФ, дЦМФ и дАМФ, причём на 5'-конце данного олигонуклеотида находится остаток дТМФ, на 3'-конце – остаток дАМФ.
Изучение ДНК методом рентгеноструктурного анализа выявило, что ее молекулы имеют пространственную структуру в виде спирали, включающей более одной полинуклеотидной цепи, а методом кислотно-щелоч-ного титрования было показано, что нативная структура ДНК стабилизируется водородными связями. Учитывая эти данные, а также основываясь на правилах Чаргаффа, Д. Уотсон и Ф. Крик в 1953 г. разработали модель строения ДНК.
Согласно модели Д. Уотсона и Ф. Крика молекула ДНК представляет собой двойную правозакрученную спираль, состоящую из двух полинуклеотидных цепей, которые закручены между собой и вокруг общей оси. Эти цепи антипараллельны и поэтому на каждом из концов двойной спирали находится 5'-конец одной цепи и 3'-конец другой. Сахарофосфатные группировки, входящие в состав нуклеотидных остатков ДНК, выходят на поверхность спиралевидной структуры, тогда как остатки азотистых оснований находятся внутри и соединяются водородными связями. При этом водородные связи возникают между парами оснований, которые структурно совместимы в пространстве. Такой принцип построения молекулы ДНК получил название комплементарного строения.
В соответствии с принципом комплементарности одна полинуклеотидная цепь двойной спирали ДНК имеет последовательность соединения нуклеотидных остатков, которая закономерно отражает другую, антипараллельную полинуклеотидную цепь (рис. 3.7). Методом построения молекулярных моделей было выяснено, что пространственно совместимыми (комплементарными) являются пары азотистых оснований аденин–тимин и гуанин–цитозин, между которыми и возникают водородные связи, удерживающие вместе в двойной спирали две полинуклеотидные цепи. При этом между остатками аденина и тимина возникают две водородные связи, а остатками гуанина и цитозина – три ( рис. 3.8).
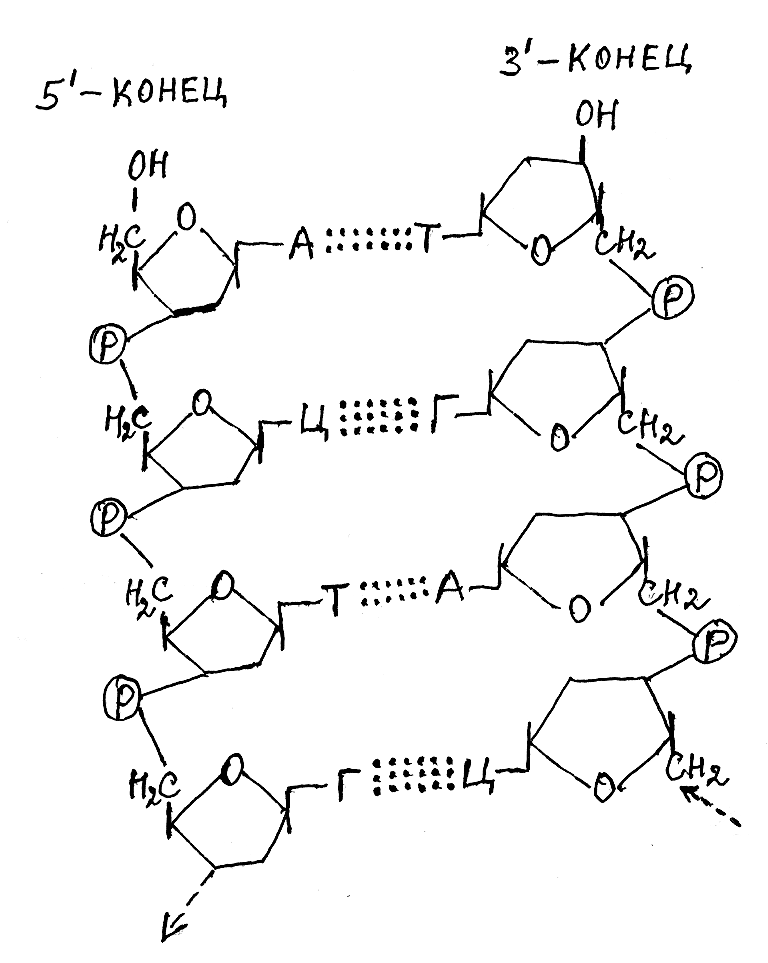
Рис. 3.7. Схема строения фрагмента ДНК

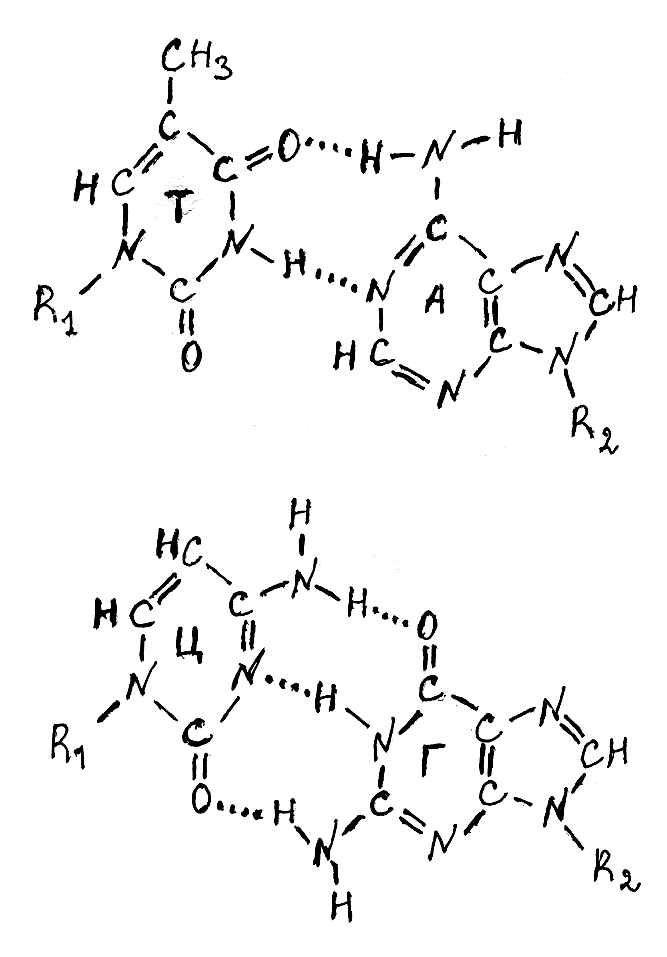
Рис. 3.8. Образование водородных связей между комплементарными основаниями в молекулах ДНК
А – аденин; Т – тимин; Ц – цитозин; R1 и R2 – комплементарные цепи двойной спирали ДНК.
Один виток спирали включает по 10 нуклеотидных остатков в каждой из двух цепей, и его длина по оси спирали составляет 3,4 нм, а межну-клеотидные расстояния – 0,34 нм. Диаметр двойной спирали ДНК – 1,8 нм. Плоскости оснований в двойной спирали ДНК перпендикулярны оси спирали. Как было установлено, модель структуры ДНК Д. Уотсона и Ф. Крика отображает пространственное строение основной формы ДНК, в виде которой она существует в клетках живых организмов. Её назвали В-формой. В В-форме остаток дезоксирибозы имеет S-конформацию, а азотистые основания – анти-конформацию.
Однако в дальнейшем было выяснено, что молекула ДНК может принимать и другую конформацию. При понижении концентрации воды в физиологической среде в молекулах ДНК изменяется пространственная структура дезоксирибозы, которая принимает N-конформацию, в результате чего уменьшаются расстояния между нуклеотидными остатками (до 0,25 нм) и увеличивается диаметр двойной спирали, а число нуклеотидных пар в одном витке спирали возрастает до 11. Однако спираль остаётся правозакрученной и конформация азотистых оснований не изменяется. Двойную спираль, имеющую указанные параметры, называют А-формой ДНК. Между А- и В-формами ДНК существуют ещё и переходные варианты пространственной структуры двойной спирали.
В ходе исследований выяснено, что некоторые участки двойной спирали ДНК под воздействием внешней среды могут переходить из правозакрученной в левозакрученную форму, которая получила название Z-формы (рис. 3.9). Такой способностью обладают участки молекулы ДНК, имеющие последовательность нуклеотидных остатков с повторяющимися парами оснований (–Г–Ц–)n. Формирование Z-формы обусловлено тем, что конформации нуклеотидных остатков в чередующихся ГЦ-последователь-ностях попарно изменяются на противоположные: в остатках дезоксицитидиловой кислоты дезоксирибоза имеет S-конформацию, а азотистое основание цитозин – анти-конформацию, тогда как в остатках дезоксигуаниловой кислоты дезоксирибоза принимает N-конформацию, а гуанин – син-конформацию.
При рассмотрении строения ДНК следует также отметить, что под влиянием внешней среды молекула ДНК может переходить из одной конформации в другую, но её нуклеотидный состав и последовательность соединения в ней нуклеотидных остатков остаются неизменными, что является важным условием сохранения генетической информации, которая заложена именно в виде полинуклеотидной последовательности.

Рис. 3.9. Пространственное изображение B- и Z-формы ДНК
(темными линиями соединены остатки ортофосфорной кислоты, отмеченные темными кружками)
Для сохранения стабильности молекул ДНК важное значение имеет принцип комплементарного связывания водородными связями оснований в двойной спирали ДНК, в результате чего, зная последовательность соединения нуклеотидных остатков в одной цепи, можно определить первичную структуру другой цепи, образующей с первой двойную спираль. Так, например, если в одной из цепей двойной спирали ДНК имеется последовательность нуклеотидных остатков –А–Г–Ц–Т–Ц–Г–, то совершенно очевидно, что в другой цепи согласно принципу комплементарности оснований образуется последовательность –Т–Ц–Г–А–Г–Ц–. Указанная закономерность в построении молекул ДНК предопределяет возможность синтеза новых молекул ДНК с сохранением заданной последовательности в ней нуклеотидных остатков, что осуществляется при размножении клеток организмов.
У высших организмов молекулы ДНК локализованы главным образом в хромосомах клеточного ядра, по одной молекуле в каждой хромосоме. Поскольку длина полинуклеотидной цепи ДНК во много раз (10 тыс. раз) превышает линейные размеры хромосом, она образует строго упорядоченные компактные структуры, которые принимают наименьшие линейные размеры в метафазных хромосомах. Основные структурные единицы при упаковке ДНК в хромосомах – нуклеосомы, которые формируются с участием гистоновых белков.
Нуклеосома представляет собой октамер, содержащий по 2 молекулы каждого из четырёх гистонов – Н2А, Н2В, Н3 и Н4, на который закручена двойная спираль ДНК длиной 150–200 пар нуклеотидных остатков. Диаметр нуклеосомы – 10 нм, толщина – 5,7 нм. Образование нуклеосомы происходит в результате взаимодействия положительно заряженных аминокислотных остатков гистонов с отрицательно заряженными фосфатными группировками нуклеотидных остатков ДНК, а также взаимодействия между гидрофобными группировками молекул гистонов, образующих гидрофобные глобулы.
Между нуклеосомами находятся участки ДНК длиной 20–90 пар нуклеотидных остатков в виде двойной спирали. Под воздействием физиологической среды нуклеосомы, соединённые свободными участками двойной спирали ДНК, формируют микрофибриллы диаметром 10 нм, которые с участием гистонов Н1 и в присутствии катионов Са2+ закручиваются в левую суперспираль с диаметром 30 нм и превращаются в крупные хроматиновые фибриллы. В свою очередь эти структуры взаимодействуют со структурными белками хромосомы и образуют суперспирализованные петли, называемые доменами, закреплённые в хромосоме в строго определённом месте. В среднем в составе одного домена обычно насчитывается 40–50 тыс. пар нуклеотидных остатков, образующих группу функционально связанных генов.
В клетках прокариот (низших организмов) двойные спирали молекул ДНК образуют замкнутую кольцевую структуру, основой формирования которой служат белки негистоновой природы. Эта структура получила название нуклеоида. В составе нуклеоида белки негистоновой природы образуют комплексы с молекулой ДНК, переводя её в высокоупорядоченное состояние.
У растений и других высших организмов не вся ДНК содержится в хромосомах ядра, часть её находится в виде кольцевых структур в составе митохондрий и пластид. Митохондриальная и пластидная ДНК содержит генетическую информацию, необходимую для осуществления этими органеллами клетки их биологических функций. Как и ДНК прокариот, кольцевые молекулы митохондриальной и пластидной ДНК при взаимодействии со специфическими белками образуют нуклеоиды.