

3.3.12. Процессы распада нуклеиновых кислот и нуклеотидов
В клетках организмов постоянно происходит расщепление полинуклеотидов на олиго- и мононуклеотиды с участием ферментов, называемых нуклеазами. Нуклеазы катализируют гидролиз фосфодиэфирных связей в молекулах РНК и ДНК, а также их фрагментов. В зависимости от расщепляемого субстрата эти ферменты называют рибонуклеазами или дезоксирибонуклеазами.
Нуклеазы различаются по специфичности действия и образуемым продуктам. Известны ферменты этого типа, обладающие невысокой специфичностью. Они способны катализировать гидролиз фосфоэфирных связей в одноцепочечных фрагментах РНК и ДНК с образованием олиго- и мононуклеотидов, имеющих на 5'-конце фосфатные группировки.
Неспецифические рибонуклеазы могут действовать на двухцепочечные участки РНК с образованием 5'- или 3'-фосфорилированных олигонуклеотидов. Неспецифические дезоксирибонуклеазы гидролитически расщепляют двухцепочечные ДНК на олиго- или мононуклеотиды, имеющие фосфатные группировки на 5'- или 3'-конце.
Специфические нуклеазы осуществляют направленный гидролиз полинуклеотидов, распознавая при своём действии или определённые нуклеотидные радикалы, или специфические последовательности нуклеотидных остатков в молекулах РНК или ДНК. Из специфических дезоксирибонуклеаз наиболее хорошо изучены рестриктазы, которые распознают в молекулах ДНК специфические последовательности из 4–6 нуклеотидных остатков. Они расщепляют двухцепочечные молекулы ДНК на строго определённые фрагменты.
Под действием набора разных нуклеаз нуклеиновые кислоты гидролизуются до свободных нуклеотидов, которые далее подвергаются распаду с участием гидролитических и окислительных ферментов.
С
нуклеазы
нуклеотидазы
нуклеиновые
кислоты
нуклеотиды
нуклеозиды
Н2О
Н2О нуклеозидфосфорилазы
азотистые
основания + рибозо-1-фосфат
Н3РО4
(дезоксирибозо-1-фосфат)
нуклеазы
нуклеотидазы
нуклеиновые
кислоты ¾¾¾®
нуклеотиды ¾¾¾®
нуклеозиды ¾®
Н2О
Н2О нуклеозидфосфорилазы
¾¾¾¾¾¾¾®
азотистые основания + рибозо-1-фосфат
Н3РО4
(дезоксирибозо-1-фосфат)
Фосфорилированные пентозы включаются в углеводный обмен, а азотистые основания могут включаться в два типа биохимических процессов – вновь вступать в синтез нуклеотидов или подвергаться дальнейшему распаду.
П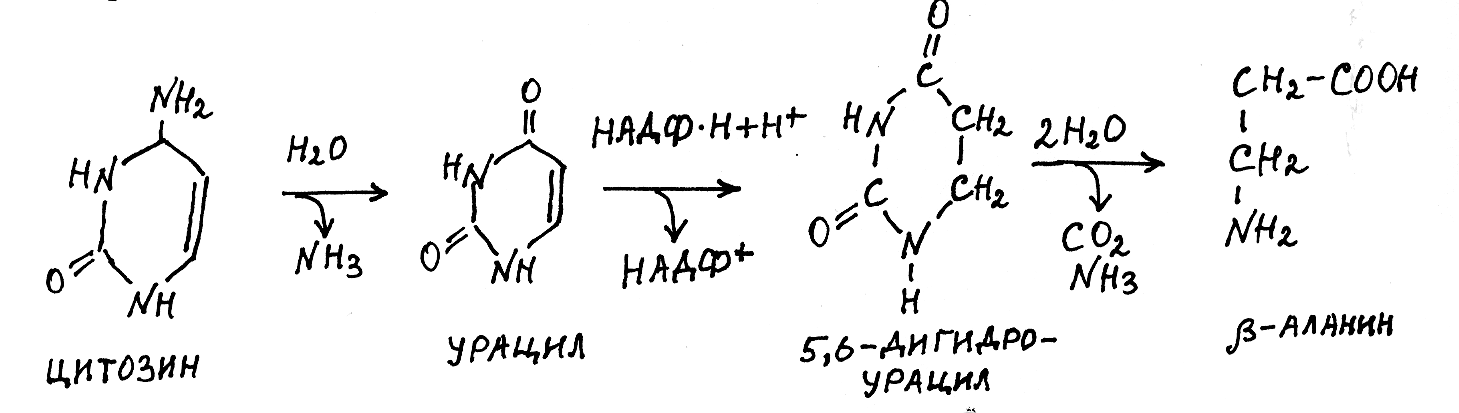 од
действием ферментов фосфорибозилтрансфераз
все азотистые основания, кроме тимина,
способны взаимодействовать с
фосфорибозилпирофосфатом и снова
включаться в рибонуклеотиды, которые
далее используются для синтеза нуклеиновых
кислот. В ходе распада азотистых оснований
цитозин в результате гидролитического
дезаминирования превращается в урацил,
который восстанавливается НАДФ
Н в 5,6-дигид-роурацил, а последний
подвергается гидролитическому расщеплению
с образованием -аланина,
СО2 и NH3:
од
действием ферментов фосфорибозилтрансфераз
все азотистые основания, кроме тимина,
способны взаимодействовать с
фосфорибозилпирофосфатом и снова
включаться в рибонуклеотиды, которые
далее используются для синтеза нуклеиновых
кислот. В ходе распада азотистых оснований
цитозин в результате гидролитического
дезаминирования превращается в урацил,
который восстанавливается НАДФ
Н в 5,6-дигид-роурацил, а последний
подвергается гидролитическому расщеплению
с образованием -аланина,
СО2 и NH3:
цитозин урацил 5,6-дигидроурацил -аланин
В свою очередь, -аланин может подвергаться окислительному расщеплению и превращаться в малонил-КоА, который при декарбоксилировании даёт ацетил-КоА. А возможные пути метаболизма малонил-КоА и ацетил-КоА нам уже известны.
Тимин в процессе распада подвергается таким же превращениям, как и урацил, только вместо -аланина образуется -аминоизобутират, который окислительным путём превращается в метилмалонил-КоА и далее в сукцинил-КоА, а последний включается в дыхательные реакции.
H2N–CH2–CH–COOH
HOOC–CH–COS–КоА
НООС–СН2–СН2–COS–КоА
|
| сукцинил-КоА
CH3
CH3
-аминоизобутират метилмалонил-КоА
П уриновые
основания аденин и гуанин, подвергаясь
гидролитическому дезаминированию,
превращаются соответственно в гипоксантин
и ксантин. Металлосодержащий флавиновый
фермент ксантиноксидаза (1.2.3.2)
окисляет гипоксантин в ксантин и далее
ксантин в мочевую кислоту. Коферментом
в молекуле этого фермента является ФАД,
ассоциированный с одним атомом молибдена
и четырьмя атомами железа.
уриновые
основания аденин и гуанин, подвергаясь
гидролитическому дезаминированию,
превращаются соответственно в гипоксантин
и ксантин. Металлосодержащий флавиновый
фермент ксантиноксидаза (1.2.3.2)
окисляет гипоксантин в ксантин и далее
ксантин в мочевую кислоту. Коферментом
в молекуле этого фермента является ФАД,
ассоциированный с одним атомом молибдена
и четырьмя атомами железа.
гипоксантин ксантин мочевая кислота
У человека мочевая кислота является
конечным продуктом распада пуринов,
который выводится из организма. Однако
у многих других организмов, включая
растения, мочевая кислота окисляется
в аллантоин. Эту реакцию катализирует
медьсодержащий фермент уратоксидаза
(1.7.3.3).
человека мочевая кислота является
конечным продуктом распада пуринов,
который выводится из организма. Однако
у многих других организмов, включая
растения, мочевая кислота окисляется
в аллантоин. Эту реакцию катализирует
медьсодержащий фермент уратоксидаза
(1.7.3.3).
мочевая кислота аллантоин аллантоиновая мочевина глиоксило-
кислота вая кислота
Аллантион в значительном количестве содержится в проростках, молодых побегах и пасоке некоторых растений. С участием фермента аллантоиназы (3.5.2.5) аллантоин гидролизуется с образованием аллантоиновой кислоты, которая также широко распространена в растениях. В клетках растений, плесневых грибов и многих рыб аллантоиновая кислота гидролитически разлагается на мочевину и глиоксиловую кислоту. Эту реакцию катализирует фермент аллантоиказа (3.5.3.4).
Таким образом, пуриновые основания в растительных клетках в результате гидролитических и окислительных реакций так же, как и в орнитиновом цикле, превращаются в мочевину, которая может затем использоваться в метаболизме азотистых веществ по уже известным нам механизмам. Следует отметить, что при высоком содержании в пище человека или кормах животных нуклеиновых кислот в результате диссимиляции пуриновых нуклеотидов образуется много мочевой кислоты, которая у этих организмов является конечным продуктом распада пуриновых оснований. При этом создаётся опасность возникновения мочекаменных заболеваний, вызванных отложением натриевых и кальциевых солей мочевой кислоты.