

65. Нервная модель стимула. Нейрофизиологические механизмы внимания.
Внимание – когнитивный процесс. Упорядочивание поступающей извне информации в аспекте приоритетности стоящих перед субъектом задач.
Внимание — это способность мозга ограничить круг объектов и явлений, с которыми человек имеет дело в процессе познания и интеллектуальной деятельности, для повышения эффективности взаимодействия с ними и улучшения качества переработки поступающей информации.
Внимание – способность мозга (человека, животного) выбирать одну информацию из потока внешних и внутренних сигналов и игнорировать другую.
Т.е., это процесс модуляции сенсорной сферы, который обеспечивает ограничение восприятия с одновременным повышением качества этого восприятия.
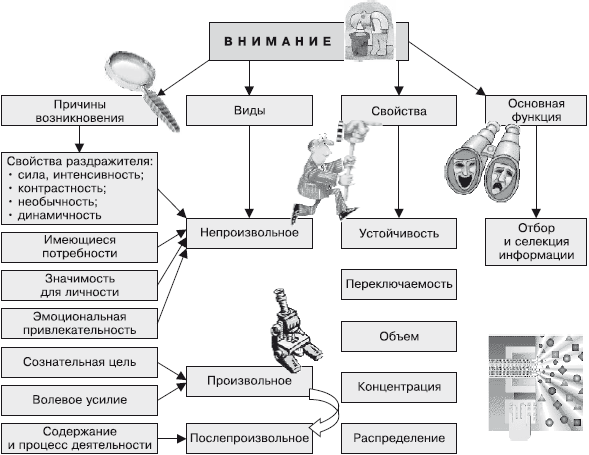
Характеристики и свойства:
Концентрация внимания — удержание внимания на каком-либо объекте. Интенсивность сосредоточения сознания на объекте.
Объём внимания — это количество однородных предметов, которые могут восприниматься одновременно и с одинаковой четкостью.
Переключаемость - сознательное и осмысленное, преднамеренное и целенаправленное, обусловленное постановкой новой цели, изменение направления сознания с одного предмета на другой.
Распределение - пособность выдерживать в центре внимания несколько разнородных объектов.
Нервная модель стимула. Механизм возникновения и угашения ОР получил толкование в концепции нервной модели стимула, предложенной Е.Н. Соколовым.
Согласно этой концепции, в результате повторения стимула в нервной системе формируется «модель» - определенная конфигурация следа, в которой фиксируются все параметры стимула.
Ориентировочная реакция возникает, когда есть рассогласование между действующим стимулом и сформированным следом, т.е. "нервной моделью".
Если действующий стимул и нервный след, оставленный предшествующим раздражителем, идентичны, то ОР не возникает. Если же они не совпадают, то ориентировочная реакция возникает и оказывается до известной степени тем сильнее, чем больше различаются предшествующий и новый раздражители.
Поскольку ОР возникает в результате рассогласования афферентного раздражения с "нервной моделью" ожидаемого стимула, очевидно, что ОР будет длиться до тех пор, пока существует эта разница.
Значимость стимула. Ориентировочный рефлекс связан с адаптацией организма к меняющимся условиям среды, поэтому для него справедлив «закон силы» -чем больше изменяется стимул (его интенсивность или степень новизны), тем значительнее ответная реакция.
Однако не меньшую, а нередко и большую реакцию могут вызвать ничтожные изменения ситуации, если они прямо адресованы к основным потребностям человека.
Кажется, что более значимый и, следовательно, в чем-то уже знакомый человеку стимул должен при прочих равных условиях вызывать меньшую ОР, чем абсолютно новый. Факты, однако, говорят о другом. Значимость стимула нередко имеет решающее значение для возникновения ОР. Высоко значимый стимул может вызвать мощную ориентировочную реакцию, имея небольшую физическую интенсивность.
Факторы, провоцирующие ОР, можно упорядочить, выделив 4 уровня, или регистра: 1) стимульный регистр; 2) регистр новизны; 3) регистр интенсивности; 4) регистр значимости.
Первый уровень оценки проходят практически все стимулы, второй и третий регистры работают параллельно. Пройдя любой из этих двух регистров, стимул поступает в последний и там оценивается его значимость. Только после этого завершающего акта оценивания развивается весь комплекс ориентировочной реакции.
Таким образом, ОР возникает не на любой новый стимул, а только на такой, который предварительно оценивается как биологически значимый. Иначе мы переживали бы ОР ежесекундно, так как новые раздражители действуют на нас постоянно.
Оценивая ОР, следовательно, надо учитывать не формальное количество информации, содержащейся в стимуле, а количество семантической, значимой информации.
Существенно и другое: восприятие значимого стимула нередко сопровождается формированием ответной адекватной реакции. Присутствие моторных компонентов свидетельствует о том, что ОР предоставляет собой единство воспринимающих и исполнительных механизмов.
Таким образом, ОР, традиционно рассматриваемая как реакция на новый раздражитель, представляет частный случай ориентировочной деятельности, которая понимается как организация новых видов деятельности, формирование активности в изменившихся условиях среды.
Нейрофизиологические механизмы внимания
Ретикулярная формация наряду с лимбической системой образуют блок модулирующих систем мозга, основной функцией которых является регуляция функциональных состояний организма.
Первоначально к неспецифической системе мозга относили в основном лишь сетевидные образования ствола мозга и их главной задачей считали диффузную генерализованную активацию коры больших полушарий.
По современным представлениям, восходящая неспецифическая активирующая система простирается от продолговатого мозга до зрительного бугра (таламуса).
Таламус часто называют «воротами в кору», поскольку, за исключением обонятельного, все сенсорные тракты проходят через него на пути к коре больших полушарий мозга.
Таламокортикальной системе отводится особая роль в обеспечении механизма селекции информации. Г. Джаспер показал, что активность неспецифических ядер таламуса вызывает реакцию десинхронизации, нарушающую медленноволновую активность. Неспецифическая система таламуса порождает локальную активацию коры за счет проекций к ее конкретным областям, воспринимающим сенсорные сигналы от модально-специфических путей.
В отличие от ретикулярной формации ствола мозга, ответственной за генерализованную активацию, захватывающую обширные зоны коры и предопределяющую ее общее функциональное состояние, таламокортикальная система обеспечивает селективное внимание и возникновение ориентировочного рефлекса.
Современные методы оценки кровенаполнения различных участков мозга помогли выявить увеличение кровотока в области ретикулярной формации и таламуса при выполнении испытуемыми заданий, требующих внимания.
В таламусе обнаружены пейсмекеры низкочастотных ритмов, и было доказано, что одни таламические структуры могут вызывать синхронизированные и ритмические колебания на ЭЭГ, другие, напротив, провоцируют реакцию активации.
Деятельность интраламинарных ядер таламуса вызывает 20-40-герцовые ритмы в коре во время активного бодрствования и 7-10-герцовые — во время второй стадии сна, когда появляются сонные веретена (Steriade e. а., 1993).
Раздражение ретикулярных ядер таламуса ведет к генерации гамма-активности в коре (Pinault, Dischenes, 1992).
Т.е. есть группы ядер, образующих «систему активации (десинхронизации)» и «систему торможения (синхронизации)».
Эти системы могут выполнять функции «фильтров», предполагаемых психологическими теориями внимания.
За селективное внимание ответственны также базальные ганглии.
Суворов и О. П. Таиров (1985) утверждают, что каждому виду деятельности соответствует особое внимание, которое может быть описано как специфическое распределение активации по коре головного мозга.
Базальные ганглии участвуют в распределении активности в коре не прямо, а опосредованно — через таламус, который выполняет функции фильтра для информации, идущей от них к коре больших полушарий.
Лимбическая система также активно участвует в изменении направления внимания, поскольку включена в формирование приоритетной информации.
Важное значение в процессе внимания придается сингулярной коре, активность которой, по-видимому, непосредственно связана с повышением значимости того или иного объекта, что проявляется в сдвиге внимания с одного явления на другое (Cochran, 1997).
Внимание ограничивает поток сенсорных сигналов за счет изменения их порога восприятия. По-видимому, кора больших полушарий мозга модулирует количество поступающей к ней информации посредством обратных связей (Coquery, 1978).
Холинергические магноцеллюлярные нейроны базального переднего мозга могут отвечать за избирательное внимание к значимым стимулам. Влияя на кору непосредственно и через таламические проекции, они модулируют ее активность в зависимости от значимости входящего сигнала.
Роль левого и правого полушарий мозга в процессе внимания.
В обеспечении мобилизационной готовности человека ведущую роль отводят правому полушарию головного мозга (Nass, Koch, 1987; Ladavas e. a., 1989).
Однако, если процесс подготовки к восприятию значимого стимула направляется выбором из нескольких возможных альтернатив с учетом предшествующей информации, то в таком случае независимо от вида информации более успешно действует левое полушарие (Cohen, 1975).
В этом полушарии в период ожидания значимого стимула складываются функциональные объединения областей, специфичные по отношению к модальности ожидаемого стимула.
Напротив, в правом полушарии характер функционального объединения областей совершенно не зависит от того, тактильный, слуховой или зрительный сигнал готовится различать испытуемый.
Если в левом полушарии связанные с направленным вниманием изменения фазовых отношений альфа-колебаний затрагивают у каждого испытуемого ограниченный круг областей, то в правом полушарии эти изменения более генерализованы и распространяются преимущественно на ассоциативные зоны (Мачинская и др., 1992).
Больные с повреждениями правого полушария имеют нарушения внимания и восприятия. Они с трудом ориентируются в пространстве, из чего сделано заключение о связи правого полушария с пространственной интеграцией внимания (Pardo, 1991).
Эксперименты с рассеченным полушарием мозга покрывает, что процессы внимания тесно связаны с работой мозолистого тела, при этом левое полушарие обеспечивает селективное внимание, а правое - поддержку общего уровня настороженности.
Методы изучения и диагностики внимания
Экспериментальное изучение физиологических коррелятов и механизмов внимания осуществляется на разных уровнях, начиная от нервной клетки и кончая биоэлектрической активностью мозга в целом. Каждый из этих уровней исследования формирует свои представления о физиологических основах внимания.