

64. Современные нейрофизиологические теории сознания (теории б.Дж.Баарса, к.Коха, с.Гринфилда).
В основе этих теорий лежат представления, согласно которым каждый стимул и каждое внутреннее переживание связаны с согласованной активацией некоторой распределенной группы нейронов, причем в каждую группу одновременно входят нейроны из различных удаленных друг от друга областей мозга.
Каждому стимулу, каждому впечатлению, каждому внутреннему состоянию сознания соответствует некоторое уникальное объединение нейронов. Каждый отдельный нейрон может в разное время входить в разные объединения. (В терминологии Коха эти объединения называются коалициями, в терминологии Гринфилда – ансамблями). Время жизни каждого такого объединения – не более несколько десятых долей секунды.
– Теория гностических нейронов – целостное восприятие каждого объекта обеспечивается активностью совершенно определенных нейронов (детекторов высшего порядка). Детекторы очень высокого порядка действительно обнаружены в мозге (например, нейроны, реагирующие на лица и на другие категории объектов), однако маловероятно, чтобы они действительно самостоятельно отвечали за формирование целостного представления об объекте (включая ассоциации в памяти, эмоциональную окраску и т.п.).
– Теория распределенного кодирования, согласно которой нейроны, занятые обработкой различных аспектов одного объекта, синхронизируют свою активность друг с другом, и в то же время разряжаются несинхронно с остальными нейронами, не участвующими в обработке информации о данном объекте. В последние 10-15 лет накопилось множество фактов, указывающих на важность синхронизированной активности (прежде всего в диапазоне гамма-ритма около 40 Гц, но также и в других частотных диапазонах).
Вероятно, первое наблюдение этого явления было сделано Вольфом Зингером в конце 80-х гг. (1989 г.), который показал, что нейроны зрительной коры, удаленные друг от друга и отвечающие за разные участки зрительного поля, синхронизировались друг с другом в том случае, если соответствующие участки зрительного поля представляли собой единый объект (две полоски на экране двигались как единое целое); синхронизация не возникала, если те же полоски двигались независимо друг от друга.
МОДЕЛЬ КОХА. Объединение пирамидных нейронов, соединяющее переднюю и заднюю области коры больших полушарий, разряжается уникальным образом. Разные объединения соответствуют различным стимулам.
МОДЕЛЬ ГРИНФИЛД. Нейроны из разных областей мозга разряжаются синхронно и доминируют до тех пор, пока другой стимул не вызывает появление нового ансамбля. Ежемоментно под влиянием входящих сигналов образуются и распадаются различные ансамбли.
Чем объясняется субъективная разница между сном и бодрствованием?
Кох: Во время фазы сна с быстрыми движениями глаз закономерности рас- пределения активности по областям мозга сильно отличаются от того, что наблюдается при бодрствовании. В частности, лимбическая система (система эмоций и памяти) очень активна, однако области лобных долей, участвующие в рациональном мышлении, подавлены. Объединения нейронов возникают как во время сна, так и во время бодрствования, однако они включают в себя нейроны из разных частей мозга. Во время бодрствования объединения могут включать значительно больше нейронов в префронтальной коре, где на восприятие накладывается логика и причинно-следственные связи, однако такая активность резко снижается во время сна. Именно этим объясняется причудливость и выраженная эмоциональная окрашенность сновидений.
Гринфилд: Сновидения связаны с ансамблями нейронов значительно меньшего размера, чем во время бодрствования. Размер ансамблей ограничен, т.к. нет сильных сенсорных стимулов, привлекающих большие коли- чества нейронов. Следовательно, кратковременное вовлечение нейронов во время сна обусловлено только лишь реакциями на собственную спонтан- ную мозговую активность. И поскольку ансамбли не запускаются естествен- ной по-следовательностью событий внешнего мира, то взаимосвязи между сменяющими друг друга ансамблями делаются хаотичными или вообще отсут-ствуют, в результате сновидения становятся случайным набором образов или событий. Отсутствие обширных действующих нервных связей также объясняет очевидное отсутствие самоконтроля, характеризующего в норме наше бодрствующее сознание.
Кох, говорит о том что есть специализированные группы нейронов, группировка которых всегда едина и как раз они отвечают за сознание, а Гринфилд говорит о том что эти образования непостоянны и образуются согласно окружающей среде.
Теория «глобального рабочего пространства» Б.Дж.Баарса
В теории Баарса мозг рассматривается как состоящий из множества различных параллельно работающих процессоров (или модулей), каждый из которых способен решать некоторую задачу над символическими репрезентациями, поступающими им на вход.
Гибкость модулей заключается в их способности комбинироваться и образовывать новые процессоры, которые могут решать новые задачи, а также распадаться на меньшие составляющие процессоры.
Баарс рассматривает мозг как большую группу распределенных частичных процессоров, высоко специализированных систем, функционирующих большую часть времени на бессознательном уровне.
По крайней мере, некоторые из этих частичных процессов могут быть представлены на сознательном уровне, когда они организуются и образуют глобальные процессы. Глобальные процессы предоставляют сознанию информацию и образуются в ходе борьбы и кооперации частичных процессоров.
Итак, Баарс постулировал существование теоретического рабочего пространства, в котором на основе взаимодействия множества частичных процессов образуются глобальные процессы. Он постулировал, что глобальные образования в рабочем пространстве обеспечивает глобальное доминантное сообщение сознательной мысли. … Глобальное образование может рассматриваться как сложная сеть частичных процессов.
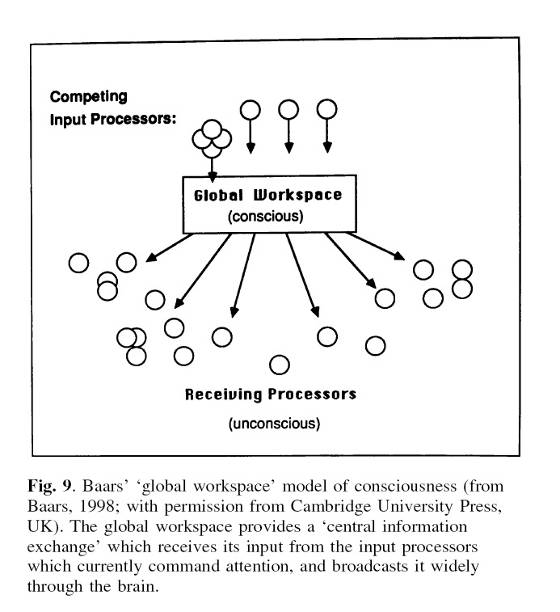
В исходной версии теории Баарса предполагалось, что образование глобального рабочего пространства связано с деятельностью неспецифических ядер таламуса и ретикулярной формации.
В настоящее время большая роль в этом процессе отводится длинным кортико-кортикальным связям, в особенности связям префронтальной коры с ассоциативной теменной корой и с поясной извилиной.