

Молекулярная основа мутаций
До сих пор мы рассматривали мутации как отдельные изменения в последовательности ДНК, влияющие на активность той генетической единицы, в которой они возникают. Если рассматривать мутации как способ инактивации генов, то у большинства генов и спонтанная, и индуцированная частота мутирования примерно одинаковы. Точнее, частота мутирования есть функция размера гена, т.е. мишени для мутаций. Но если рассматривать мутационные сайты (под сайтом в данном случае понимают одну пару оснований) внутри определенной последовательности ДНК, следует задать вопрос: все ли пары оснований в одинаковой степени чувствительны к изменениям или некоторые из них мутируют с большей частотой, чем другие?
Мутации концентрируются в горячих точках
Многочисленные генетические и биохимические исследования показали, что возникающие мутации локализуются вовсе не равномерно по всему геному. В некоторых сайтах происходит гораздо больше мутаций, чем следует ожидать при их случайном распределении. Их количество в определенных сайтах может быть в 10 и даже 100 раз больше случайного статистического возникновения. Такие сайты называют горячими точками.
Долгое время природа горячих точек была не ясна. Считали, что горячие точки должны соответствовать каким-то последовательностям, особенно чувствительным к спонтанным и индуцированным мутациям. Действительно, было обнаружено, что сайты одних типов оказывались горячими точками для спонтанных мутаций и мутаций, индуцированных одними мутагенами, тогда как другие мутации возникают в результате действия других мутагенов.
Главной причиной спонтанных точечных мутаций у E. coli является присутствие необычных модифицированных оснований, которые получили название минорных компонентов ДНК. Свое название модифицированных они получили вследствие того, что эти основания образуются посредством химической или ферментативной модификации одного из четырех обычных оснований, присутствующих в норме в ДНК. Однако в модифицированной форме, под действием ферментов репликации или репарации они в ДНК не включаются. Среди таких модифицированных оснований, которые появляются в ДНК уже после репликации наиболее известны 5-метилцитозин (5-Me-Cyt), 7-метилгуанин (7-Me-Gua) и 6-N-метиладенин (N6-Me-Ade). Наиболее распространенное модифицированное основание в составе ДНК – это 5-метилцитозин (рис. 5.2), который находят у про- и эукариот, и о котором уже упоминалось при описании структуры Z-формы ДНК. Модификацию осуществляет фермент метилаза, который добавляет метильную группу к цитозину только в определенных участках ДНК, модифицируя лишь незначительную часть всех остатков цитозина.

Рис. 5.2 |
Модификация остатков цитозина в составе молекул ДНК под действием метилазы. |
В одном наиболее детально исследованном гене (ген lac I, который входит в состав lac-оперона, см. главу «Контроль генной экспрессии у прокариот») все горячие точки спонтанных, а также индуцированных точечных мутаций попадают в сайты, которые у дикого типа содержат 5-метилцитозин. Во всех случаях мутации возникали в результате транзиции G-C в А-Т. Более того, в штаммах E. coli с нарушенными процессами метилирования горячие точки обнаружены не были.
Причина таких замен заключается в том, что при спонтанном или индуцированном дезаминировании 5-Me-Cyt образуется Thy (рис. 5.3), а это, в свою очередь, приводит к ошибочному спариванию G с Т, что при последующей репликации может вызывать появление транзиции G-C A-T.
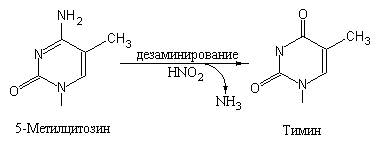
Рис. 5.3 |
Дезаминирование 5-метилцитозина приводит к появлению тимина, который не в состоянии образовать каноническую пару с G, что может вызвать транзицию G-C A-T. |
В данном контексте уместно привести существующее предположение относительно того, почему в ДНК используется азотистое основание Thy вместо Ura, который, как известно, является обычным основанием для молекул РНК. Если в результате окислительного дезаминирования Cyt в ДНК появляется Ura, он воспринимается как ненормальное для дезоксирибонуклеиновой кислоты основание и поэтому удаляется под действием репарирующего фермента урацил-ДНК-гликозилазы (см. далее). В результате в соответствующем сайте остается неспаренный остаток гуанина, и система репарации по правилам комплементарности включает в поврежденный участок ДНК утраченный цитозин. Если бы в ДНК присутствовал в качестве обычного нормального основания Ura, то в случае дезаминирования Cyt ферментные системы не могли бы распознать какой из остатков Ura является правильным, а какой появился в результате дезаминирования Cyt. При окислительном дезаминировании 5-метилцитозина образуется тимин, который является обычным компонентом ДНК. Поэтому в таких случаях система репарации не всегда оказывается способной узнавать ошибку, что приводит к появлению мутации.
Другой важной известной горячей точкой в гене lac I является последовательность CTGG, которая повторяется три раза подряд. Мутации возникают в результате делеции или вставки одной четырехбуквенной единицы. Такие мутации могут появляться в результате «проскальзывания» при репликации, если одна цепь не точно совпадает с другой. Например, первая четырехбуквенная единица одной цепи может спариться со второй четырехбуквенной единицей другой цепи. В этом случае может произойти добавление или потеря четырехбуквенных единиц.
Мутант CTGG CTGG
делеция
Дикий тип CTGG CTGG CTGG
вставка