

Молекулярная основа мутаций. Причины мутаций. Действие физических факторов
Одним из фундаментальных требований, предъявляемых к структуре, которая выполняет роль постоянного хранилища информации является ее чрезвычайная стабильность. Эта стабильность существенна, по крайней мере, в смысле тех характеристик структуры, которые обеспечивают кодирование генетической информации. Поэтому необходимым свойством структуры ДНК является высокая стабильность содержания в ней азотистых оснований и их последовательности. Однако, целостность оснований, их состав и последовательность в молекулах ДНК не полностью ограждены от постепенных изменений. Обычно такие изменения происходят не часто и затрагивают всего несколько оснований. Различные химические и физические факторы могут модифицировать структуру определенных оснований или даже разрушать фосфодиэфирные связи, приводя тем самым, к разрывам цепей. Ошибки также могут возникать в процессе репликации или рекомбинации, вызывая встраивание одного или нескольких дополнительных оснований в новую цепь. Вместе с тем, почти в любом случае необходимо осуществление нескольких циклов репликации для того, чтобы появившаяся модификация могла привести к необратимому изменению в ДНК. Другими словами, ДНК-полимераза должна использовать в качестве матрицы изначально поврежденную полинуклеотидную цепь для того, чтобы исходные изменения стали постоянными (см. рис. 5.1). Как показано на рис. 5.1, использование в качестве матрицы поврежденной цепи обеспечивает распространение повреждения от замены одного единственного основания в одной цепи до полного изменения пары оснований в обеих цепях, а последующая репликация навсегда сохранит это изменение. Необратимые изменения в структуре ДНК получили название мутаций.
Открытие генетической роли ДНК естественным образом породило представление о том, что мутации возникают в результате изменений в последовательности нуклеотидов.
Некоторые мутации возникают в результате нормальных, обычных процессов в клетке, или при взаимодействии нуклеинового материала с окружающей средой. Такие мутации называют самопроизвольными или спонтанными мутациями: они появляются с определенной частотой у любого организма.
Однако, частоту мутаций можно существенно увеличить, воздействуя определенными соединениями. Наследственные изменения, возникающие под действием таких соединений называют индуцированными мутациями, а сами соединения - мутагенами. Большинство мутагенов действует прямо, реагируя либо с определенными азотистыми основаниями, либо встраиваясь в нуклеиновую кислоту.
В процессе мутагенеза любая пара оснований может мутировать. Изменения, которые затрагивают только одну пару оснований, называют точечными мутациями. Точечные мутации могут быть двух типов. К первому типу точечных мутаций относят замены одного пурина на другой пурин, или одного пиримидина на другой пиримидин и называют такие мутации транзициями. Примером транзиции может служить замена пары C-G на T-A или наоборот. Это наиболее часто встречающийся тип точечных мутаций. Ко второму типу точечных мутаций относят трансверсии, которые характеризуются заменами пурина пиримидином и наоборот, т.е. пара A-T превращается в T-A или C-G.
Одной из возможных причин точечных мутаций, относящихся к транзициям является химическое превращение одного основания в другое. Примером может служить превращение цитозина (Сyt) в урацил (Ura) либо спонтанное, в ходе обычной реакции окислительного дезаминирования Сyt, либо под действием, например, азотистой кислоты (HNO2). Как показано на рис. 5.1, после дезаминирования Сyt, в последующем цикле репликации образовавшийся Ura спаривается с аденином (Ade) вместо гуанина (Gua), с которым спаривался бы исходный Сyt. Таким образом, пара C-G замещается парой A-T, что совершенно понятно, так как в последующем цикле репликации в пару с Ade становится тимин (Thy). В свою очередь, при действии азотистой кислоты может также происходить дезаминирование Ade, вызывая обратную транзицию A-T в G- C.
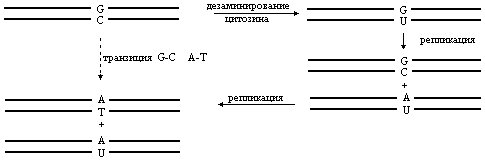
Рис. 5.1 |
Схема, иллюстрирующая последовательность событий приводящих к возникновению транзиции G-C A-T. |
Другой возможной причиной транзиций может быть ошибочное спаривание оснований, приводящее к образованию неканонических пар и, следовательно, к искажению двойной спирали ДНК. При обычной репликации такая ошибка может случайно произойти вследствие включения неправильного основания, представляющего собой редкую таутомерную форму. В этом случае спонтанная частота ошибок определяется прежде всего степенью точности функционирования ДНК-полимеразы. Существует также «репаративный» синтез ДНК, который активируется в результате генетической рекомбинации или возникших повреждений ДНК. Различные системы репарации также характеризуются разной частотой ошибок и, таким образом, могут даже увеличивать частоту наследуемых изменений в последовательности ДНК.
Некоторые соединения, которые можно отнести к мутагенам представляют собой аналоги обычных природных оснований, но эти аналоги способны к неканоническому спариванию. В процессе репликации они могут включаться в ДНК. Из-за ошибочного спаривания при их включении или при последующей репликации они могут вызвать целую серию транзиций. Примером такого мутагена может служить 5-бромурацил (5-Br-Ura). Это соединение включается в ДНК вместо тимина, но из-за того, что 5-Br-Ura достаточно эффективно спаривается также с гуанином (вместо аденина), его присутствие ведет к образованию замен пар А-Т на G-C. В каждом цикле репликации эти замены возникают с определенной вероятностью, поэтому 5-Br-Ura, однажды включенный в ДНК вместо тимина, продолжает индуцировать транзиции в последующих циклах репликации.
К следующему классу мутагенов относятся акридиновые соединения. Их связывание с ДНК настолько деформирует двойную спираль, что это приводит к вставкам (инсерциям) дополнительных оснований или к выпадениям (делециям) оснований при репликации. В результате каждого мутагенного акта, индуцированного акридином, исчезают или добавляются одна или несколько пар оснований. Мутации такого типа называют мутациями со сдвигом рамки считывания.
Долгое время считали, что основным типом мутаций в индивидуальных генах являются точечные мутации. Однако, в настоящее время известно, что, по крайней мере, у бактерий очень часто происходят вставки (инсерции) дополнительного материала или выпадение (делеции) части или целой вставки, а иногда и соседних с вставками участков ДНК. Источником этих вставок служат подвижные генетические элементы – определенные последовательности ДНК, которые способны перемещаться из одного места хромосомы в другое. Следует иметь в виду, что вставки и делеции могут возникать (хотя и с меньшей вероятностью) и по другим причинам – например, в результате ошибок во время репликации или рекомбинации.