

Строение синтазы жирных кислот
Биосинтез ЖК осуществляется мультиферментным комплексом, который называется синтазой жирных кислот (пальмитатсинтаза, СЖК). Строение этого комплекса приведено на рис.23.1.
Мультиэнзимный комплекс синтазы ЖК у высокоорганизованных форм (млекопитающие, птицы, насекомые) характеризуется молекулярной массой 400-500 кДа, образован единственной полипептидной цепи, включающей около 2300 аминокислотных остатков. Этот комплекс состоит из двух идентичных субъединиц, т.е. является гомодимером. Каждая субъединица может катализировать семь различных реакций, из которых складывается синтез ЖК. Пространственное объединение нескольких реакций в мультиферментный комплекс (МФК) имеет ряд преимуществ по сравнению с отдельными ферментами: предотвращаются конкурентные реакции, последовательные реакции согласованы как на конвейере, реакции протекают эффективно, благодаря высокой концентрации субстрата и незначительным потерям его за счет диффузии.
У бактерий, таких, как E. Coli в синтезе ЖК участвуют семь отдельных ферментов и ацилпереносящий белок. Растения также имеют индивидуальные белки с различной активностью, которые ассоциированы в четвертичный комплекс.
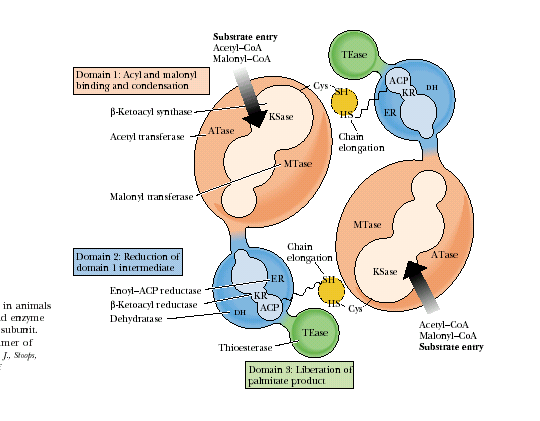
Рис.23.1. Комплекс синтазы жирных кислот млекопитающих
Каждая субъединица МФК включает три различных домена и восемь субдоменов.
Домен I состоит из трех субдоменов: субдомен 1 − АПБ-S-ацетилтрансфераза, 60 кДа; субдомен 2 − АПБ-S-малонилтрансфераза, 23 кДа; субдомен 3 − β-кетоацил-АПБ-синтаза (конденсирующий фермент, 45 кДа). Домен I катализирует присоединение субстратов ацетил-СоА и малонил-СоА ацетилтрансферазой и малонилтрансферазой соответственно и последующую конденсацию обоих партнеров β-кетоацил-синтазой.
Домен II также состоит из трех субдоменов: субдомен 4 − β-кетоацил-АПБ-редуктаза, 21 кДа; субдомен 5 − β-гидроксиацил-АПБ-дегидратаза, 50 кДа; субдомен 6 − еноил-АПБ-редуктаза, 14 кДа. К субдомену 4 присоединен ацилпереносящий белок (АПБ), 15 кДа. Домен II восстанавливает растущую цепь ЖК с помощью вышеназванных трех ферментов.
Домен III содержит субдомен 7 − ацил-АПБ-гидролаза, тиоэстераза, 33 кДа. Домен III после семь циклов удлинения цепи катализирует высвобождение готового продукта − пальмитата с помощью гидролитического фермента тиоэстеразы.
Центральную роль в функционировании комплекса СЖК играет ацилпереносящий белок, простетической группой которого является 4-фосфопантетеин. Почти посередине полипептидной цепи этого белка находится остаток серина, к ОН-группе которого сложноэфирной связью присоединяется 4-фосфопантетеин, по структуре идентичный фосфопантотеиновой группе коэнзима А (рис.23.2).

Рис.23.2. Фосфопантетеиновая группа коэнзима А и ацилпереносящего белка
Функция АПБ в биосинтезе ЖК аналогична функции СоА в β-окислении жирных кислот. В процессе наращивания цепи ЖК промежуточные продукты связаны с АПБ. Простетическая группа АПБ служит «вращающейся ручкой», которая перемещает промежуточные соединения от активного центра одного фермента к другому. Ацильные остатки прочно связываются с концевой SH-группой АПБ, что предотвращает обмен метаболитами, образующимися в процессе синтеза и распада ЖК. Кроме HS-группы АПБ в синтазе жирных кислот есть еще одна SH-группа, принадлежащая специфическому остатку цистеина в молекуле β-кетоацил-синтазы. Обе эти SH-группы участвуют в биосинтезе жирных кислот. Растущая цепь ЖК всегда связана с фосфопантетеиновой группой ацилпереносящего белка.
Димер синтазы жирных кислот ориентирован таким образом, что Домен I одного мономера прилежит к Доменам II и III другого мономера. Поворот «фосфопантетеиновой ручки» позволяет реакции активации протекать на одной субъединице, а остальным реакциям цикла − на другой. Поэтому синтаза ЖК функционально активна только в виде димера.