

Терминация транскрипции
Последовательности ДНК, являющиеся сигналами к остановке транскрипции, называются транскрипционными терминаторами. Они содержат инвертированные повторы (GC-богатые участки), благодаря чему 3ʹ-концы РНК-транскриптов складываются с образованием шпилек разной длины. Поскольку сила взаимодействия пар G-C велика, локальная денатурация таких участков в ДНК затруднена. Это замедляет продвижение РНК-полимеразы и может служить для нее сигналом к прекращению транскрипции.
Обнаружены два типа сигналов терминации: ρ-зависимый и ρ-независимый терминаторы (ρ ‒ это олигомерный белок, прочно связывающийся с РНК и в этом состоянии гидролизующий АТР до АDP и неорганического фосфата).
В одной из моделей действие ρ-белка объясняется тем, что он связывается с синтезируемой цепью РНК и перемещается вдоль нее в направлении 5ʹ→ 3ʹ к месту синтеза РНК, необходимая для перемещения энергия выделяется при гидролизе АТР (рис.30.6).

Рис.30.6. Механизм ρ-зависимой терминации транскрипции
При этом ρ-белок наталкивается на образующуюся в РНК «шпильку» и останавливает полимеразу, которая могла бы продолжить транскрипцию. Механизм ρ-независимой терминации изучен хуже, в нем остается много неясного.
Транскрипция у эукариот
РНК-полимеразы эукариот изучены значительно слабее, чем соответствующие ферменты бактерий. Синтез РНК у эукариот осуществляют три различных фермента: РНК-полимеразы I, II и III. Их структура изучена не полностью. РНК-полимераза I необходима для синтеза 18S-, 5,8S- и 28S-рибосомальных рНК; РНК-полимераза II участвует в синтезе мРНК, некоторых мяРНК; РНК-полимераза III необходима для синтеза 5S рРНК, тРНК и некоторых мяРНК. У эукариот ни одна из РНК-полимераз не способна самостоятельно связываться с промоторами транскрибируемых ими генов. Для этого необходимы специфичные для каждой РНК-полимеразы белковые факторы транскрипции (TF-факторы) (рис. 30.7). Механизмы транскрипции у эукариот и прокариот идентичны.
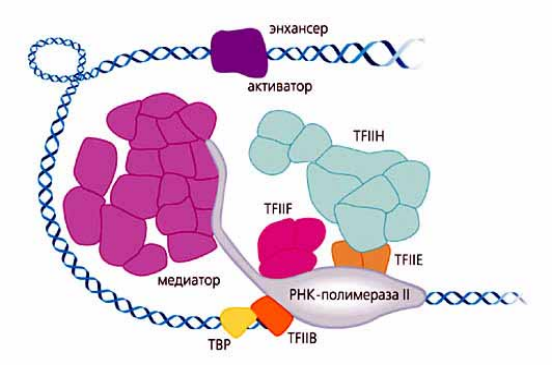
Рис.30.7. Комплекс инициации транскрипции у эукариот
На рис. 30.7 в составе комплекса приведены общие факторы транскрипции (TFIIB, E, F, H и TBP), РНК-полимераза II, медиатор и специфический фактор транскрипции, связанный с энхансером
Процессинг первичных транскриптов РНК
В большинстве случаев первичные транскрипты, образующиеся вышеписанным способом, не являются зрелыми молекулами РНК, а требуют процесса созревания, который называется процессингом РНК. У прокариот первичные транскрипты, сформированные при транскрипции генов, кодирующих белки, функционируют в качестве мРНК без последующей модификации или процессинга. Более того, трансляция мРНК часто начинается даже до завершения синтеза 3ʹ-конца транскрипта. Совсем иная ситуация наблюдается для молекул прокариотических рРНК и тРНК. В этом случае кластеры рРНК- или т-РНК-генов часто транскрибируются с образованием единой цепи РНК. Для формирования зрелых функциональных форм необходимо специфическое разрезание первичных РНК-транскриптов и их модификация. Эти молекулярные события и называются процессингом РНК (или посттранскрипционной модификацией).
Созревание первичных транскриптов у эукариот осуществляется гораздо сложнее. Первичные транскрипты, так называемые гетерогенные ядерные РНК (гяРНК), по размерам значительно превышают цитоплазматические мРНК, участвующие в трансляции. Это связано с тем, что гяРНК содержат интроны. На первом этапе созревания интроны должны быть удалены из гяРНК. Этот процесс называется сплайсингом (от англ. to splice ‒ сплетать, сращивать). После удаления интронов происходит ковалентное соединение участков РНК, которые были транскрибированы с экзонов (рис.30.8).

Рис.30.8. Схема сплайсинга РНК
Сплайсинг катализируется нуклеопротеиновым комплексом, сплайсосомой, в состав которой, наряду с белками, входят малые ядерные РНК (мяРНК). Это небольшие по размеру молекулы, характерной особенностью которых является значительное содержание пиримидинового основания ‒ урацила (рис.30.9).

Рис.30.9. Структура сплайсосомы
В первичных транскриптах именно мяРНК катализируют расщепление фосфодиэфирных связей. Именно они являются рибозимами. В сплайсосомах место сплайсинга определяется с высокой точностью, поскольку ошибка даже в один нуклеотид может привести к синтезу функционально неактивного белка. Точное узнавание осуществляется благодаря специфическим нуклеотидным последовательностям (сигналам) на концах интронов.
Кроме сплайсинга, мРНК эукариот подвергается модификации: на 5ʹ-конце синтезируется «кэп» (шапочка) ‒ структура, представляющая собой метилированный остаток гуанозинтрифосфата, который защищает РНК от гидролиза 5ʹ-экзонуклеазами. На 3ʹ-конце пре-мРНК синтезируется полиаденилатная последовательность длиной 150-200 нуклеотидов − «шлейф», поли-А-хвост (см. лекцию 11).
Процессинг прешественников тРНК и рРНК у эукариот осуществляется аналогично таковому у прокариот (рис.30.10)

Рис.30.10. Процессинг пре-транспортной РНК
Регуляция генной экспрессии на уровне транскрипции
Клетки имеют механизмы, регулирующие синтез различных белков. Размер хромосомной ДНК в Е.соli (4,7∙106 нуклеотидных пар) достаточен для кодирования примерно 4000 белков. Однако единовременно синтезируется лишь около 1000 белков. В 46 хромосомах человека число закодированных белков в 100 раз больше, но не все белки постоянно синтезируются. Это означает то, что в клетке биосинтез белков контролируется, причем некоторые белки синтезируются только при определенных условий.
Белки, синтезируемые с постоянной скоростью, называются конститутивными, белки, синтезируемые со скоростью, резко изменяющейся в зависимости от условий – адаптивными, или индуцибельными.
Конститутивные белки (в том числе и ферменты) содержатся в клетках примерно в постоянном количестве, независимо от того, есть ли в них потребность. Предполагается, что они будут нужны клетке всегда. Поэтому промоторы кодирующих их генов не имеют «выключателя», они всегда находятся в положении «включено». Конститутивными белками являются, например ферменты гликолиза. Количество молекул адаптивных белков варьируется в широких пределах. Очевидно, что синтез конститутивных белков не регулируется, а индуцибельных, напротив, подвергается тонкой регуляции.
Стимуляция биосинтеза белков, сопровождающаяся увеличением их количества, называется индукцией, подавление синтеза белков – репрессией. В клетках имеются вещества, сигнализирующие о состоянии метаболизма внутри клеток или организма. С помощью этих веществ можно «включать» или «выключать» синтез белков. Такими веществами у прокариот могут быть поступающие в клетку питательные вещества и некоторые внутриклеточные регуляторы (циклические нуклеотиды).
Впервые схема регуляции биосинтеза белков у прокариот была предложена французскими учеными Ф. Жакобом и Ж. Моно в 1961 году. Эта схема была разработана на примере лактозного оперона Е.соli и получила название механизма индукции – репрессии активности генов. Их идеи были подтверждены в ходе различных генетических и биохимических исследований.
Механизм регуляции активности разных транскриптонов (оперонов) у бактерий выглядит следующим образом. В бактериях имеется группа белков, называемых репрессорами, которые контролируют транскрипцию разных оперонов. Участок ДНК, определяющий структуру белка-репрессора, называется геном-регулятором. Он может быть расположен рядом с опероном, а может располагаться на определенном расстоянии от регулируемого оперона.
Все репрессоры связываются с оператором оперона и блокируют транскрипцию; в этом случае не синтезируются определенные мРНК и не идет синтез соответствующих белков.
Способность связываться с оператором зависит от конформации репрессора, которая может быть активной и неактивной. Только в активной форме репрессор способен образовывать слабые связи с оператором и блокировать синтез мРНК и белка; в неактивной форме он не может соединяться с оператором. Вещества, которые инактивируют репрессор, называются индукторами (рис. 30.11).

Рис.30.11.Регуляция экспрессии генов путем индукции
Вещества, переводящие его из неактивного состояния в активное, называются корепрессорами. Следовательно, репрессор имеет участок связывания корепрессора и индуктора (рис.30.12)

Рис.30.12. Регуляция экспрессии генов пути репрессии