

Геномные мутации
Геномные мутации, как и хромосомные, являются причинами возникновения хромосомных болезней.
К геномным мутациям относятся анеуплоидии и изменения плоидности структурно неизмененных хромосом. Геномные мутации выявляются цитогенетическими методами.
Анеуплоидия – изменение (уменьшение – моносомия, увеличение – трисомия) числа хромосом в диплоидном наборе, некратное гаплоидному (2n+1, 2n-1 и т.д.).
Полиплоидия – увеличение числа наборов хромосом, кратное гаплоидному (3n, 4n, 5n и т.д.).
У человека полиплоидия, а также большинство анеуплоидий являются летальными мутациями.
К наиболее частым геномным мутациям относятся:
трисомия – наличие трех гомологичных хромосом в кариотипе (например, по 21-й паре при болезни Дауна, по 18-й паре при синдроме Эдвардса, по 13-й паре при синдроме Патау; по половым хромосомам: XXX, XXY, XYY);
моносомия – наличие только одной из двух гомологических хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона не возможно. Единственная моносомия у человека, совместимая с жизнью – моносомия по Х-хромосоме – приводит к синдрому Шерешевского-Тернера (45,Х).
Причиной, приводящей к анеуплодии, является нерасхождение хромосом во время клеточного деления при образовании половых клеток или утрата хромосом в результате анафазного отставания, когда во время движения к полюсу одна из гомологичных хромосом может отстать от других негомологичных хромосом. Термин нерасхождение означает отсутствие разделения хромосом или хроматид в мейозе или митозе.
Нерасхождение хромосом наиболее часто наблюдается во время мейоза. Хромосомы, которые в норме должны делиться во время мейоза, остаются соединенными вместе и в анафазе отходят к одному полюсу клетки, таким образом, возникают две гаметы, одна из которых имеет добавочную хромосому, а другая – не имеет этой хромосомы. При оплодотворении гаметы с нормальным набором хромосом гаметой с лишней хромосомой возникает трисомия (т.е. в клетке присутствует три гомологичные хромосомы), при оплодотворении гаметой без одной хромосомы возникает зигота с моносомией. Если моносомная зигота образуется по какой-либо аутосомной хромосоме, то развитие организма прекращается на самых ранних стадиях развития.
Мутации — это стойкие внезапно возникшие изменения структуры наследственного материала на различных уровнях его организации, приводящие к изменению тех или иных признаков организма. Термин «мутация» введен в науку Де Фризом. Им же создана мутационная теория, основные положения которой не утратили своего значения по сей день.
Мутации возникают внезапно, скачкообразно, без всяких переходов.
Мутации наследственны, т.е. стойко передаются из поколения в поколение.
Мутации не образуют непрерывных рядов, не группируются вокруг среднего типа (как при модификационной изменчивости), они являются качественными изменениями.
Мутации ненаправленны — мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков в любом направлении.
Одни и те же мутации могут возникать повторно.
Мутации индивидуальны, то есть возникают у отдельных особей.
Процесс возникновения мутаций называют мутагенезом, а факторы среды, вызывающие появление мутаций, — мутагенами.
По типу клеток, в которых мутации произошли, различают: генеративные и соматические мутации.
Генеративные мутации возникают в половых клетках, не влияют на признаки данного организма, проявляются только в следующем поколении.
Соматические мутации возникают в соматических клетках, проявляются у данного организма и не передаются потомству при половом размножении. Сохранить соматические мутации можно только путем бесполого размножения (прежде всего вегетативного). По адаптивному значению выделяют: полезные, вредные (летальные, полулетальные) и нейтральные мутации. Полезные — повышают жизнеспособность, летальные — вызывают гибель, полулетальные — снижают жизнеспособность, нейтральные — не влияют на жизнеспособность особей. Следует отметить, что одна и та же мутация в одних условиях может быть полезной, а в других — вредной. По характеру проявления мутации могут быть доминантными и рецессивными. Если доминантная мутация является вредной, то она может вызвать гибель ее обладателя на ранних этапах онтогенеза. Рецессивные мутации не проявляются у гетерозигот, поэтому длительное время сохраняются в популяции в «скрытом» состоянии и образуют резерв наследственной изменчивости. При изменении условий среды обитания носители таких мутаций могут получить преимущество в борьбе за существование. В зависимости от того, выявлен ли мутаген, вызвавший данную мутацию, или нет, различают индуцированные и спонтанные мутации. Обычно спонтанные мутации возникают естественным путем, индуцированные — вызываются искусственно. В зависимости от уровня наследственного материала, на котором произошла мутация, выделяют: генные, хромосомные и геномные мутации.
Закон гомологических рядов в наследственной изменчивости Н.И. Вавилов, изучая наследственную изменчивость у культурных растений и их предков, обнаружил ряд закономерностей, которые позволили сформулировать закон гомологических рядов наследственной изменчивости: «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство». Этот закон можно проиллюстрировать на примере семейства Мятликовые, к которому относятся пшеница, рожь, ячмень, овес, просо и т.д. Так, черная окраска зерновки обнаружена у ржи, пшеницы, ячменя, кукурузы и других растений, удлиненная форма зерновки — у всех изученных видов семейства. Закон гомологических рядов в наследственной изменчивости позволили самому Н.И.Вавилову найти ряд форм ржи, ранее не известных, опираясь на наличие этих признаков у пшеницы. К ним относятся: остистые и безостые колосья, зерновки красной, белой, черной и фиолетовой окраски, мучнистое и стекловидное зерно и т.д. Открытый Н.И.Вавиловым закон справедлив не только для растений, но и для животных. Так, альбинизм встречается не только в разных группах млекопитающих, но и птиц, и других животных. Короткопалость наблюдается у человека, крупного рогатого скота, овец, собак, птиц, отсутствие перьев у птиц, чешуи у рыб, шерсти у млекопитающих и т.д. Закон гомологических рядов наследственной изменчивости имеет огромное значение для селекционной практики. Он позволяет предугадать наличие форм, не обнаруженных у данного вида, но характерного для близкородственных видов, то есть закон указывает направление поисков. Причем искомая форма может быть обнаружена в дикой природе или получена путем искусственного мутагенеза. Например, в 1927 г. немецкий генетик Э.Баур, исходя из закона гомологических рядов, высказал предположение о возможном существовании безалкалоидной формы люпина, которую можно было бы использовать на корм животным. Однако такие формы не были известны. Было высказано предположение, что безалкалоидные мутанты менее устойчивы к вредителям, чем растения горького люпина, и большая их часть погибает еще до цветения. Опираясь на эти предположения, Р.Зенгбуш начал поиски безалкалоидных мутантов. Он исследовал 2,5 млн. растений люпина и выявил среди них 5 растений с низким содержанием алкалоидов, которые явились родоначальниками кормового люпина. Более поздние исследования показали действие закона гомологических рядов на уровне изменчивости морфологических, физиологических и биохимических признаков самых разных организмов — от бактерий до человека.
Фенокопии и генокопии
Довольно часто при реализации разных генотипов могут возникать сходные фенотипы: фенокопии и генокопии.
Термин «фенокопия» употребляется в том случае, если рассматриваются «дикий» и мутантный генотипы. Корректное применение термина «фенокопия» предполагает, что для одного генотипа данный результат считается нормальным, а для другого – аномальным. Фенокопии – это, в сущности, морфозы и тераты. Они не наследуются, но наследуетсяпредрасположенность к образованию фенокопий.
Пример 1. У насекомых темная окраска тела может быть обусловлена генетически. Однако при низких температурах появляются меланистические формы и у насекомых с генотипом, который при в стандартных условиях дает обычную окраску. Тогда морфоз «темное тело» является фенокопией мутации «темное тело».
Пример 2. Яровой тип развития у злаков обусловлен определенным генотипом. Яровизация озимых злаков обусловлена воздействием внешних факторов. Тогда развитие озимых злаков по яровому типу является фенокопией наследственно яровых форм.
Примеры фенокопий у дрозофилы (по И.А. Рапопорту):
Мутация (генотип) |
Химические индукторы морфозов у мушек «дикого типа» (генотип +/+) |
Фенотип (фенокопия мутантного фенотипа) |
yellow (генотип самок y/y, генотип самцов y/Y) |
азотнокислое серебро |
желтое тело |
brown (bw/ bw) |
соединения сурьмы |
коричневые глаза |
Minute |
соединения ртути |
тонкие щетинки |
eyeless (ey/ey) |
соединения бора |
безглазие |
aristopedia |
соединения бора |
превращение арист в ноги (ариста – придаток последнего членика усика) |
Термин «генокопия» используется, если рассматривается два и более мутантных генотипа. Например, у дрозофилы ярко-красную окраску глаз обеспечивают мутации в разных генах: v, cn, st, cd. Тогда дрозофилы с разными генотипами, но ярко-красными глазами будут генокопиями друг друга.
Механизмы возникновения генокопий различны. Например, цепочка превращений исходного вещества в конечный продукт X→Y→Z может быть прервана в результате мутаций в гене A, контролирующем переход X→Y, или в гене B, контролирующем переход Y→Z.
Часто мутагенные факторы называют мутагенами (от мутации и греческого genes - рождающий, рождённый). Мутагены, увеличивающие частоту мутаций в сотни раз (нитропроизводные мочевины) называются супермутагенами.
Процесс образования мутаций с помощью физических или химических мутагенов называется мутагенезом. Последний является одним из важнейших приёмов экспериментальной генетики. Часто термины "мутагенез" и "мутационный процесс" отождествляются, что не оправдано, т.к. мутационный процесс - это многоэтапный процесс возникновения спонтанных или индуцированных мутаций, а мутагенез - это процесс индукции мутаций.
Радиоактивным мутагенезом начали заниматься в 20-х годах нашего столетия. В 1925 г. советские учёные Г.С. Филиппов и Г.А. Надсон впервые в истории генетики применили рентгеновские лучи для получения мутаций у дрожжей. Через год американский исследователь Г. Меилер (впоследствии дважды лауреат Нобелевской премии) применил тот же мутаген к мушке дрозофиле.
Химический мутагенез впервые целенаправленно начал изучать В.В. Сахаров в 1931 г. на дрозофиле при воздействии на её яйца йодом.
Мутации называют прямыми, если их проявление приводит к отклонению признаков от так называемого дикого типа, наиболее распространённого в природе, и обратными (реверсиями), если их проявление приводит к полному или частичному восстановлению дикого типа.
Мутации бывают:
* генеративными (происходят в половых клетках и в этом случае передаются последующим поколениям);
* соматическими (происходят в любых других - соматических - клетках организма и в этом случае наследуются только при вегетативном размножении);
* ядерными (затрагивают хромосомы ядра);
* цитоплазматическими (затрагивают генетический материал, заключённый в цитоплазматических органоидах клетки - митохондриях, пластидах и т.п.).
В зависимости от характера изменений в генетическом материале различают следующие мутации: точковые, инсерции, хромосомные перестройки или аберрации, и мутации, заключающиеся в изменении числа хромосом.
Точковые мутации представляют собой вставки или выпадения, а также изменения (транзиции и трансверсии) пары нуклеотидов ДНК (или нуклеотида РНК). Они могут изменять функцию отдельных генов, а также нескольких соседних генов одного оперона в случае полярного эффекта, т.е. выключения всех генов, расположенных дистально от оператора по отношению к возникшей мутации (полярные мутации).
Инсерции - вставки молекул ДНК в ген, приводящие чаще всего к его инактивации или сильному полярному эффекту в оперонах.
Хромосомные перестройки возникают в основном, по-видимому, за счёт "незаконной" рекомбинации, т.е. рекомбинации негомологичных участков генетического материала. К ним относятся: делеции, в том числе дефишенси (концевые нехватки хромосом); инверсии, дупликации, транслокации, транспозиции - перемещения участков генетического материала, соизмеримых по длине с геном, между хромосомами или в пределах одной хромосомы.
Изменения числа хромосом в клетках организма могут быть кратными гаплоидному набору (анеуплоидия).
В зависимости от возникновения выделяют три главные группы мутаций: генные, хромосомные и геномные.
К генным относят все точковые мутации, к геномным - изменения числа хромосом.
Хромосомные аберрации могут быть отнесены как к генным, так и к хромосомным мутациям, в зависимости от размера участка, затрагиваемого аберрацией.
Если рассмотреть эти группы мутаций (генные, хромосомные и геномные) более подробно, то становится ясно следующее: мутировать может часть гена или весь ген целиком, благодаря чему ген может существовать в виде двух или многих аллелей. В опытах с дрозофилой получено множество генных мутаций, касающихся цвета глаз, тела, формы крыльев и других признаков.
Однако, мутировать могут и более крупные единицы - части хромосом, целые хромосомы или даже геномы. Среди хромосомных мутаций встречаются два основных типа: межхромосомные мутации (транслокации) и структурные изменения хромосом (инверсии, делеции и дупликации). Примером хромосомной мутации может служить дупликация гена Bar - плосковидные глаза Ultra Bar у дрозофилы.
Последняя группа - геномные мутации, при некоторых происходит изменение числа хромосом. Они могут быть двух типов: гетероплоидии и полиплоидии.
Гетероплоидия заключается в изменении числа какой-либо (или каких-нибудь) части генома. В результате гетероплоидии, явления болезненного или даже губительного, образуются особи с ненормальным (вместо обычного диплоидного - 2n) числом хромосом: моносомия 2n-1, трисомия 2n+1, полисомии (2n+2, 2n+3) и т.п.
Полиплоидия - наличие в клетках увеличенного в кратное число раз гаплоидного набора. Полиплоидия может быть естественной и искусственной и придавать возникающим особям новые свойства и качества. Среди животных как в естественных, так и в искусственных условиях, полиплоидия наблюдается сравнительно редко. Зато в мире растений полиплоидия широко распространена. Около трети видов растений, произрастающих на нашей планете - полиплоиды. Почти все культурные растения, в том числе и цветы, украшающие наши сады, тоже полиплоиды. В эволюции растений полиплоидия являлась как стимулятором, так и тормозом прогресса. Многие древние растения, например, древовидные папоротники, магнолии и др., были полиплоидами.
Мутации могут изменять поведение, коснуться любых физиологических особенностей и затрагивать строение особи. Большинство изученных мутаций касается именно изменения морфологических признаков. Существуют биохимические мутации, которые выражаются в изменении какого-либо фермента.
По влиянию на жизнеспособность особи мутации могут быть летальными, полулетальными, сублетальными, снижающими в большей или меньшей степени жизнеспособность организма. Большинство из мутаций вредны для организма, нередко мутантный ген может обусловить наследственное заболевание, уродство или даже гибель развивающегося организма. Могут быть мутации практически нейтральные в данных условиях, прямо не влияющие на жизнеспособность. Очень редко возникают генные мутации, улучшающие те или иные свойства (полезные мутации). Именно они дают основной материал для естественного и искусственного отбора, являясь необходимым условием эволюции в природе и селекции полезных форм растений, животных и микроорганизмов.
Мутагенез — процесс возникновения наследственных изменений организма — мутаций. Редкое появление среди нормальных организмов измененных особей было известно давно. Однако научное описание явлений мутагенеза было сделано лишь в 1899 г. рус. ученым С. И. Коржинским и в 1900—1901 гг. голландским генетиком Г. де Фризом, который, в частности, ввел термины «мутация» (лат. изменение) и «мутагенез». Способность мутировать присуща всем формам жизни на Земле и лежит в основе фундаментального свойства живого — изменчивости. Сущность мутаций состоит в изменении структуры дезоксирибонуклеиновой кислоты (ДНК), являющейся носителем генетической информации (см. Нуклеиновые кислоты). Мутации приводят к возникновению нового признака или изменению (утрате) старого. Различают спонтанный (возникающий в естественных условиях без четко регистрируемых внешних воздействий) и индуцированный (в результате искусственных воздействий) мутагенеза.
Природа спонтанного мутагенеза изучена недостаточно. Долгое время ученые считали, что спонтанные мутации возникают под действием естественного фона радиации (в т. ч. космических лучей), а также остаточных количеств некоторых химических веществ, образующихся за счет разложения природных органических соединений или в процессе хозяйственной и бытовой деятельности человека. Одним из факторов спонтанного мутагенеза являются химические соединения, естественно образующиеся в организме как промежуточные продукты обмена веществ. Многие ученые считают ведущей причиной спонтанного мутагенеза «ошибки» ферментов, взаимодействующих с ДНК при размножении клеток, устранении повреждений ДНК (см. Репарация генетическая) или в процессе обмена генами (см. Ген). Экспериментально установлено, что частота спонтанного мутагенеза близка к частоте «ошибок» ферментов, участвующих в генетических процессах. Индуцированные мутации возникают под влиянием повреждающего действия на генетический аппарат клеток некоторых фиэических и химических агентов. Впервыеиндуцированные (искусственные) мутации были получены в 1925 г. советскими учеными Г. А. Надсоном и Г. С. Филипповым путем облучения клеток дрожжей рентгеновскими лучами. В 1927 г. американский генетик Г. Меллер установил мутагенное действие рентгеновских лучей в отношении мушки дрозофилы. В 1928 г. советский микробиолог М. Н. Мейсель опубликовал результаты исследований, доказавших возможность получения мутаций при воздействии на клетки химических соединений — так называемый химический мутагенез, широкое изучение которого стало проводиться с начала 30-х гг. в основном советскими генетиками (В. В. Сахаров, С. М. Гершензон и др.). Теоретические основы этого направления разрабатывались советским генетиком И. А. Рапопортом, английским ученым Ш. Ауэрбахом, американским ученым Э. Фризом. Факторы, вызывающие мутации (различные виды излучений, химических соединений), стали называть мутагенами.
Мутагены взаимодействуют с молекулами ДНК, непосредственно изменяя их структуру, или оказывают на них опосредованное действие.
Прямое действие на ДНК сопровождается, как правило, пропорциональным возрастанием частоты мутаций при увеличении дозы или времени воздействия мутагена. Однако довольно часто зависимость мутагенеза от дозы облучения, времени обработки или от концентрации химических мутагенов оказывается более сложной, что обусловлено в основном двумя группами причин.
Во-первых, при взаимодействии мутагенов с ДНК (или с хромосомой как морфология, структурой) в ряде случаев для появления мутаций однократного воздействия мутагенного фактора оказывается недостаточно; мутация возникает лишь после двух или большего числа воздействий. Во-вторых, как уже говорилось, мутагены могут взаимодействовать не прямо с ДНК, а оказывать опосредованное (непрямое) действие. Напр., мутаген может вызывать образование в клетках значительного количества различных промежуточных химических соединений (радикалов), которые в дальнейшем взаимодействуют с хромосомами и ДНК, изменяя их строение. Многие мутагены сохраняются в клетках длительное время (нередко в течение нескольких клеточных делений), проявляя так называемые последействие. Кроме того, установлено, что нередко повреждения, возникающие в ДНК, остаются в неизменном состоянии в течение длительного времени, тогда как проявление этих повреждений в виде новых мутаций имеет место у далеких потомков особей, подвергшихся мутагенному воздействию.
Изменение скорости и характера мутирования происходит также из-за вмешательства в мутагенез различных ферментов, способных устранять из ДНК ее поврежденные звенья (см. Репарация генетическая). Таким образом, появление мутаций, как правило, не является следствием простого взаимодействия мутагенов с наследственным веществом клеток. возникновении генных мутаций одно или несколько оснований в структуре гена могут перемещаться на другое место в этой же или в другие хромосомы (транслокации), удваиваться (дупликации), теряться (делеции) или оставаться на том же месте в хромосоме, но оказываться повернутыми на 180° (инверсии) и др. Хромосомные мутации подразделяют на дупликации, при которых отдельные участки хромосом-удваиваются или утраиваются; инверсии, когда участки хромосом оказываются перевернутыми на 180° транслокации, при которых наблюдается перенос участков хромосом на другое место в пределах той же хромосомы или в другую хромосому: делеции, сопровождающиеся утратой частей хромосом большей или меньшей длины; фрагментации, при которых хромосома распадается на значительное число участков и прекращает свое существование в виде морфологически целостной структуры; и циклизации, приводящие к замыканию концов хромосомы и образованию кольцевой хромосомы.
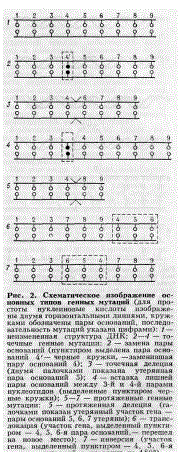
В случае геномных мутаций весь хромосомный набор может или уменьшаться вдвое (гаплоиды) или увеличиваться вдвое и больше раз (полиплоиды). Могут быть также потеряны отдельные -хромосомы набора (анеуплоиды).ности ферментов генетической репарации, которые полностью или частично устраняют повреждения, видоизменяют структуру ДНК, по возможности возвращая ее молекулу к исходному нормальному строению. Все это сказывается на скорости мутагенеза. Типы мутаций. Мутации подразделяют на три типа: генные, хромосомные и геномные (рис. 1). Изменения, затрагивающие структуру ДНК в пределах отдельных генов, называют генными мутациями. В свою очередь, генные мутации подразделяют на точечные и протяженные (рис. 2). Под точечными мутациями понимают изменения одиночных азотистых оснований в структуре ДНК: замена одних оснований другими, приводящая к изменению генетической записи (см. Генетический код) в пределах данного гена; выпадения или вставки отдельных звеньев ДНК (нуклеотидов), нарушающие порядок считывания генетической информации в пределах гена таким образом, что вся информация вслед за измененной точкой будет прочитана неправильно. Естественным следствием описанных мутаций будет изменение структуры кодируемого мутировавшим геном белка и соответствующего наследственного признака.
 Протяженные
генные мутации подразделяют на дупликации
(участок гена удваивается или утраивается),
инверсии (происходит два разрыва ДНК в
пределах гена, и выпавший фрагмент ДНК
снова присоединяется на то же самое
место, но с поворотом на 180°), транслокации
(выпавший фрагмент присоединяется в
новом месте в пределах гена) и делеции
(возникают в том случае, если выпавший
участок гена теряется). Хромосомные
мутации (хромосомные перестройки, или
аберрации). На стадии деления клеток
каждая хромосома состоит из двух
продольных частей, называемых хроматидами.
В зависимости от того, является ли
аберрация следствием изменения одной
хроматиды или хромосомы в целом,
перестройки называют хроматидными и
хромосомными. Хромосомные перестройки
возникают после того как образуется
один или несколько разрывов хромосом
или отдельных хроматид. Оторвавшийся
участок хромосомы может нек-рое время
существовать в клетке в виде одиночного
фрагмента, но при делении клетки он
может быть потерян, и в месте отрыва
возникает хроматидная или хромосомная
делеция. Освободившиеся в результате
разрывов концы хромосом могут
взаимодействовать друг с другом, формируя
так называемые обменные аберрации, при
которых участки хромосомы могут оказаться
в другом месте в той же хромосоме или
же переместиться в другую хромосому с
сохранением порядка расположения генов
(транслокация) или поворотом выпадающего
участка на 180° (инверсия).
Протяженные
генные мутации подразделяют на дупликации
(участок гена удваивается или утраивается),
инверсии (происходит два разрыва ДНК в
пределах гена, и выпавший фрагмент ДНК
снова присоединяется на то же самое
место, но с поворотом на 180°), транслокации
(выпавший фрагмент присоединяется в
новом месте в пределах гена) и делеции
(возникают в том случае, если выпавший
участок гена теряется). Хромосомные
мутации (хромосомные перестройки, или
аберрации). На стадии деления клеток
каждая хромосома состоит из двух
продольных частей, называемых хроматидами.
В зависимости от того, является ли
аберрация следствием изменения одной
хроматиды или хромосомы в целом,
перестройки называют хроматидными и
хромосомными. Хромосомные перестройки
возникают после того как образуется
один или несколько разрывов хромосом
или отдельных хроматид. Оторвавшийся
участок хромосомы может нек-рое время
существовать в клетке в виде одиночного
фрагмента, но при делении клетки он
может быть потерян, и в месте отрыва
возникает хроматидная или хромосомная
делеция. Освободившиеся в результате
разрывов концы хромосом могут
взаимодействовать друг с другом, формируя
так называемые обменные аберрации, при
которых участки хромосомы могут оказаться
в другом месте в той же хромосоме или
же переместиться в другую хромосому с
сохранением порядка расположения генов
(транслокация) или поворотом выпадающего
участка на 180° (инверсия).
 При
нарушении в клетках структур, обеспечивающих
деление клеток (митоз), отдельные
удвоившиеся хромосомы или весь набор
хромосом после его удвоения могут
остаться в той же клетке. Такие мутации
называют геномными. В случае нерасхождения
по дочерним клеткам отдельных хромосом
возникают клетки, называемые анеуплоидными,
при нерасхождении полных наборов
хромосом возникают полиплоидные клетки.
Частным случаем этого типа мутаций
является получение организмов, в клетках
которых содержится не два, как обычно,
а лишь один набор хромосом. Такие клетки
называют гаплоидными. Мутагены.
Первоначально искусственное получение
мутаций было достигнуто с помощью
жестких излучений (рентгеновские лучи),
затем было установлено, что мутации
могут вызывать ультрафиолетовые лучи,
потоки заряженных частиц (быстрые и
медленные протоны, электроны, мезоны и
др.), нейтроны, обломки ядер атомов и
другие виды излучений и элементарных
частиц. Излучения, способные проникать
в клетки, могут ионизировать молекулы
ДНК и тем самым непосредственно изменять
гены. При этом может происходить как
изменение оснований в ДНК и их разрушение
(деструкция), так и разрыв ДНК (рис. 3, 2,
3, 5, 6). Ионизирующие излучения, кроме
того, могут приводить к образованию
различных радикалов, к-рые также
воздействуют на ДНК. При уменьшении
концентрации кислорода в клетках, как
правило, происходит уменьшение количества
образующихся радикалов и параллельно
с этим падает частота мутаций (так
называемый кислородный эффект). Уменьшение
частоты мутаций, вызываемых ионизирующими
излучениями, происходит также при
введении защитных веществ —
радиопротекторов.
При
нарушении в клетках структур, обеспечивающих
деление клеток (митоз), отдельные
удвоившиеся хромосомы или весь набор
хромосом после его удвоения могут
остаться в той же клетке. Такие мутации
называют геномными. В случае нерасхождения
по дочерним клеткам отдельных хромосом
возникают клетки, называемые анеуплоидными,
при нерасхождении полных наборов
хромосом возникают полиплоидные клетки.
Частным случаем этого типа мутаций
является получение организмов, в клетках
которых содержится не два, как обычно,
а лишь один набор хромосом. Такие клетки
называют гаплоидными. Мутагены.
Первоначально искусственное получение
мутаций было достигнуто с помощью
жестких излучений (рентгеновские лучи),
затем было установлено, что мутации
могут вызывать ультрафиолетовые лучи,
потоки заряженных частиц (быстрые и
медленные протоны, электроны, мезоны и
др.), нейтроны, обломки ядер атомов и
другие виды излучений и элементарных
частиц. Излучения, способные проникать
в клетки, могут ионизировать молекулы
ДНК и тем самым непосредственно изменять
гены. При этом может происходить как
изменение оснований в ДНК и их разрушение
(деструкция), так и разрыв ДНК (рис. 3, 2,
3, 5, 6). Ионизирующие излучения, кроме
того, могут приводить к образованию
различных радикалов, к-рые также
воздействуют на ДНК. При уменьшении
концентрации кислорода в клетках, как
правило, происходит уменьшение количества
образующихся радикалов и параллельно
с этим падает частота мутаций (так
называемый кислородный эффект). Уменьшение
частоты мутаций, вызываемых ионизирующими
излучениями, происходит также при
введении защитных веществ —
радиопротекторов.

Иное действие на генетический аппарат клеток оказывает ультрафиолетовое излучение. В довольно широком диапазоне волн эти лучи поглощаются нуклеиновыми кислотами и, воздействуя на электроны атомов, входящих в состав ДНК, и передавая им свою энергию, возбуждают их. Это повышает реакционную способность групп атомов в составе азотистых оснований в ДНК, после чего в месте возбуждения азотистые основания либо реагируют с молекулами воды, либо соединяются между собой, образуя так наз. димеры (рис. 3, 4). Иногда на значительном протяжении в ДНК разрываются связи, соединяющие две цепочки молекулы ДНК, и освободившиеся в этих участках основания получают возможность взаимодействовать с окружающими их молекулами. Наряду с радиационными описано большое число (несколько сотен) химических мутагенов, которые оказались в ряде случаев даже более мощными мутагенными факторами, чем ионизирующее и ультрафиолетовое излучение. По времени действия на генетический аппарат клеток мутагены могут быть разделены на два типа: действующие на покоящуюся ДНК и способные вызывать мутации лишь во время репликации (удвоения) ДНК. Мутагены первого типа способны присоединять к азотистым основаниям и фосфатным группам в ДНК разнообразные группировки атомов (рис. 4). После этого либо изменяются кодирующие свойства ДНК и во время ее репликации напротив измененных оснований встраиваются неправильные основания, либо происходит разрыв остова молекул ДНК, что может, в частности, приводить к возникновению хромосомных перестроек. После действия многих мутагенов часть азотистых оснований может быть утеряна и тогда на место образовавшихся брешей могут присоединиться любые основания, что опять-таки приводит к нарушению смысла генетической записи. Возникшие при этом изменения проявляются в замене аминокислот в белках, строящихся под контролем поврежденного гена.

К мутагенам второго типа относятся разнообразные соединения, близкие или аналогичные по структуре азотистым основаниям, и другие вещества, соединяющиеся с ДНК и мешающие правильной ее репликации. Некоторые из них приводят к вставке в ДНК лишних оснований или выпадению из ее молекул отдельных звеньев — нуклеотидов (так наз. мутации сдвига чтения). Наконец, известны химические мутагены, действующие главным образом во время репликации ДНК, но способные оказывать мутагенное воздействие и на покоящуюся ДНК. Воздействие таких химических соединений приводит к мощному увеличению частоты мутирования. Мутагенное действие оказывают также бактериальные вирусы, вирусы насекомых и млекопитающих, препараты ДНК. Хотя относительная частота появляющихся при их действии мутаций невелика, однако типы возникающих мутаций и специфичность мутагенеза разнообразны. Мутации и здоровье человека. Клетки организма человека подвержены мутированию как в нормальных условиях жизнедеятельности (спонтанные мутации), так и при воздействии радиационными и химич. мутагенами (индуцированный мутагенез). Частота спонтанного мутирования у человека изучена для многих признаков; ее можно сопоставить с частотой мутирования у других организмов. В среднем на 1 млн. рождений в одном случае наблюдается появление наследственной болезни, обусловленной спонтанной мутацией в определенном гене. Однако, учитывая, что у человека имеется, по крайней мере, несколько десятков тысяч генных локусов, можно подсчитать, что около 15% всех гамет (половых клеток) может нести в себе новую патологическую мутацию. Несмотря на грубую приближенность этих оценок и невозможность пока дать более точные оценки, следует признать, что эти цифры подтверждаются наблюдениями генетиков и врачей. Это означает, что при имеющемся населении Земли не менее 70 млн. новорожденных несут новые мутации. Эта огромная цифра может еще увеличиться, если в окружающей нас среде будут накапливаться вредные для наследственности вещества, если в атмосфере по-прежнему будут производиться ядерные взрывы и если человечество не примет действенных мер против загрязнения окружающей среды.
Канцерогене́з (лат. cancerogenesis; cancer — рак + др.-греч. γένεσις — зарождение, развитие) — сложный патофизиологический процесс зарождения и развития опухоли. (син. онкогенез).
Изучение процесса канцерогенеза является ключевым моментом как для понимания природы опухолей, так и для поиска новых и эффективных методов лечения онкологических заболеваний. Канцерогенез — сложный многоэтапный процесс, ведущий к глубокой опухолевой реорганизации нормальных клеток организма. Из всех предложенных до ныне теорий канцерогенеза, мутационная теория заслуживает наибольшего внимания. Согласно этой теории, опухоли являются генетическими заболеваниями, патогенетическим субстратом которых является повреждение генетического материала клетки (точечные мутации, хромосомные аберрации и т. п.). Повреждение специфических участков ДНК приводит к нарушению механизмов контроля за пролиферацией и дифференцировкой клеток и в конце концов к возникновению опухоли[1].
Генетический груз — накопление летальных и сублетальных отрицательных мутаций, вызывающих при переходе в гомозиготное состояние выраженное снижение жизнеспособности особей, или их гибель.
«Вырождение» — наблюдаемое при близкородственном скрещивании ухудшение фенотипических характеристик потомства. [1]
В более строгом смысле генетический груз в популяционной генетике — это выражение уменьшения селективной ценности для популяции по сравнению с той, которую имела бы популяция, если бы все индивидуальные организмы соответствовали бы наиболее благоприятному генотипу. Обычно выражается в среднейприспособленности по сравнению с максимальной приспособленностью.
Частью генетического груза является мутационный груз.
Генетический груз рассматривается, как мера неприспособленности популяции к условиям окружающей среды. Он оценивается по различию приспособленности реальной популяции — по отношению к приспособленности воображаемой, максимально приспособленной популяции.
Значение генетического груза обычно находится в интервале 0 < L < 1, где 0 — отсутствие генетического груза. Примерами генетического груза в человеческих популяциях являются аллели мутантных форм гемоглобина —Гемоглобина С и Гемоглобина S.
Насле́дственные заболева́ния — заболевания, возникновение и развитие которых связано с дефектами впрограммном аппарате клеток, передаваемыми по наследству через гаметы. Термин употребляется в отношенииполиэтиологических заболеваний, в отличие от более узкой группы — генные болезни.
Наследственные заболевания — болезни, обусловленные нарушениями в процессах хранения, передачи и реализации генетической информации