

Фотофосфорилювання
Зі зменшенням енергії електрону спряжене фотофосфорилювання – отже втрачена енергія електрону запасається в молекулах АТФ.
Може бути циклічне або нециклічне фосфорилювання.
Пункти спряження фотофосфорилювання знаходяться на електроннотранспортному ланцюгу.
У пунктах спряження енергія повинна перевищувати ЕАТФ
![]()
ΔG електроннотранспортного ланцюга від первинного Донора +0,82 В до кінцевого Акцептора -0,32 В.
Різниця Еred/ox 0,18 В – отже термодинамічна можливість виникає коли >= 0,18 В, а саме у 3 пунктах спряження донорно-ацкепторних пар:
Пластохінон А (0,36 В)-цитохром f (0,11В) ΔЕо=0,25В;
Цитохром b(0,06В)-пігмент Р700 (0,43В) ΔЕо=0,37В;
Н2О↔1/2О2+Н2 (0,82В)-пігмент Р680 (-0,05В) ΔЕо=0,87В
На кожні два ē утворюється дві молекули АТФ
Для відновлення СО2 до [H-CH-OH] потрібно 3 АТФ і 2 НАДФ·Н2.
Відповідно по 2 АТФ і 2 НАДФ утворюється на 4 електрони нециклічного щляху.
Фотосинтез 8 квантовий, бо
![]()
на кожен ē припадає 2 кванти світла.
ККД ФОТОСИНТЕЗУ
При засвоєнні 1М СО2 запасається в продуктах фотосинтезу 471 кДж енергії.
Середнє значення 1 Е фотосинтетичної активності реакції вартує 209,34 кДж, оскільки утворюється 8Е =1674,7 кДж:
звідси ККД становитиме 471/1674,7 *100% = 28%
ФОТОФОСФОРИЛЮВАННЯ
Енергетичне спряження
Мембрану тилакоїдів називають спрягаючою по аналогії з мембраною мітохондрій, бо в них іде і перенесення ē та водночас синтез АТФ. Отже для пояснення фотофосфорилювання можна застосувати теорію Мітчела: електрохімічний потенціал Δμ залежить від Δφ та ΔрН
Тилакоїд – Δφ становить 50 мВ, ΔрН=3,2;
Мітохондрії – Δφ становить 200 мВ, ΔрН=набагато менше.
Отже докази правомірності теорії Мітчела для хлороплатсів наявні, однак втрачається поняття пунктів спряження.
Каталітичний цикл АТФ-ази
Кожна αβ - пара на F1 має нуклеотидзв'язуючий центр.
Кожен зв'язуючий центр почергово може знаходитись у одному з 3-х конформаційних станів:
1. T-конформація (Tight) - сильний зв’язок АЦ з АТФ.
2. L-конформація (Loose) -слабий зв’язок АЦ з АДФ.
3. O-конформація (Open) –вільний АЦ.
Протон-рухова сила повертає γ-субодиницю, яка “виштовхує” АТФ і переводить АЦ у О-конформацію.
БАКТЕРІАЛЬНИЙ ФОТОСИНТЕЗ
Ціанобактерії використовують
цитохром b6f-комплекс і пластохінони, як
для фотофосфорилювання, так і для окиснювального фосфорилювання.
У обох випадках відбувається генерація електрохімічного потенціалу на мембрані, який є енергетичним джерелом
для синтезу АТФ.
У фотосинтетичних бактерій немає ФС ІІ (тобто не відбувається фотолізу молекули води), замість води – джерелом електронів є H2S, H2 іт.д.
На початку 70 років виявлено, що бактеріальні клітини можуть прямо перетворювати енергію світла у енергію АТФ (галобактерії)
ФОТОРЕГУЛЯТОРНІ СИСТЕМИ
До фоторегуляторних систем належать фітохроми, їхня дія полягає у тому, що світлозалежні процеси індукуються при 660 нм (червоне світло), але припиняються при 730 нм (довге червоне світло).
Фітохроми виділені з багатьох об'єктів, з рослин, існує в двох взаємозалежних формах
![]()
Попередником Фд.ч.с. є І безколірна форма, швидкості утворення попередників відрізняються на 1-2 порядки.
Реакції функціонування Ф поширюються на мембранні явища, з якими пов'язана поява вторинних месенджерів, яких виділяють 2 класи: 1) діють на мембрани та метаболізм; 2) на гени (гіпотеза Мора), через сигнальний ланцюг, який активує “потенційно активні гени”.
ДІЯ СВІТЛОВИХ ПРОМЕНІВ
Ультрафіолет (100-400 нм) поділяють на вакуумну (100-200 нм), далеку (200-300 нм), близьку (300-400 нм) області.
Видима (400-750 нм).
Інфрачервона (750нм-3000 мкм) поділяють на близьку (0,75-2,5 мкмнм), середню (2,5-15 мкм), далеку (15-3000 мкм) області.
ДІЯ УФ НА БІОЛОГІЧНІ СИСТЕМИ
Ультрафіолетові промені представляють важливий природній фактор, дія якого зумовлює в живих клітинах фотохімічні реакції.
У спектрі УФ променів виділяють три ділянки :
УФ-А 400-325 нм;
УФ-B 325-280 нм;
УФ-С 280-200 нм.
При прохоженні УФ через атмосферу потік сильно послаблюється, в цьому відіграє роль озоновий шар атмосфери, який поглинає промені УФ-С і тому біля поверхні землі реєструють так званий “екологічний УФ”, який представлений УФ-А та УФ-В.
В основному біологічна дія УФ променів полягає у пошкодженні біомакромолекул, а саме білків та НК. Поглинання УФ НК зумовлено π→π* переходами в системі пуринових та піримідинових основ.
Основним типом фотохімічних реакцій НК є утворення піримідинових димерів, які утворюються в результаті зшивки двох сусідніх піримідинових основ (тимін).
Інші пошкодження НК також відіграють важливу роль в мутагенезі та канцерогенезі, наприклад, розриви полінуклетидного ланцюга, що зумовлює зменшення молекулярної маси ДНК.
Це у свою чергу може призвести до внутрішньо- чи міжмолекулярних зшивок ДНК, а також формування ДНК-білок комплексів.
Пошкодження УФ променями білкових молекул зумовлене фотохімічним перетворенням тирозину, фенілаланіну та триптофану.
У результаті фотофізичних процесів утворюються катіон радикали, які можуть дисоціювати на нейтральні радикали.
Сольватовані електрони, які утворюються у процесах дисоціації володіють високою реакційною здатністю.
Фотоінактивація ферментів власне і зумовлена фотохімічними перетвореннями триптофану та цистеїну
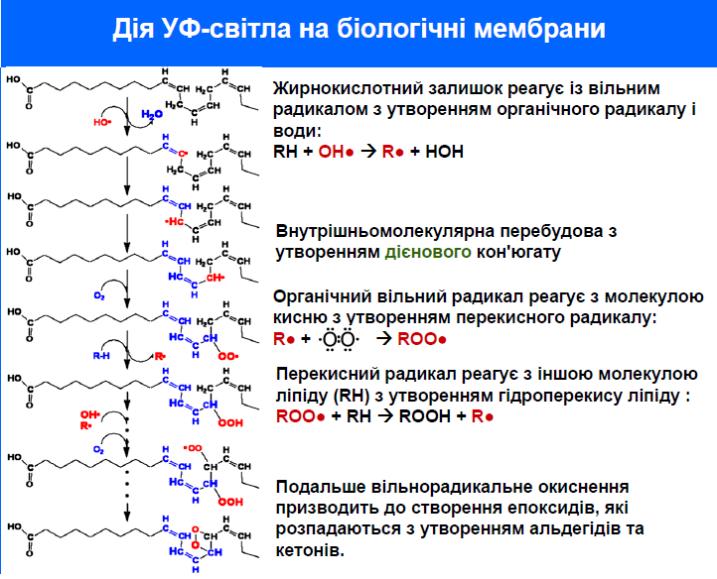
Ліпіди поглинають УФ промені в короткохвильовій ділянці. Наприклад, для ненасичених жирних кислот максимум приблизно складає 200 нм.
Основним фотохімічним ефектом дії УФ променів є окислення ліпідів. Первинним відносно стабільним продуктом фотоокиснення є гідроперекисі жирних кислот. Типовим представником кінцевих продуктів фотохімічн их перетворень є малоновий альдегід.
Перекисне окиснення ліпідів супроводжується порушенням функції мембран, оскільки з молекулярними пошкодженнями ліпідів дезорганізується структура мембран, продукти є токсичними для клітинами сполуками.
ФОТОРЕАКТИВАЦІЯ
ДНК в клітині здатна відновлюватися від основного індукованого УФ променями пошкодженнями – від піримідинових димерів. Цей процес проходить в темноті і називається темновою реактивацією.
Проходить він і на світлі шляхом фотореактивацією.
Суть полягає в тому, що під впливом видимого світла проходить розпад пряме фоторозщеплення димерів або неферментна фотореактивація.
Для клітин однак більш характерний процес ферментативної реактивації – за умов дії ферменту фотоліази (широко поширена у бактерій, дріжджів, клітин вищих рослин, тварин, людини).
ФОТОЗАХИСТ
Стійкість клітин до УФ променів підвищується якщо перед опроміненням їх піддати впливу видимого чи довгохвильового УФ світла – така дія світла називається фотозахистом і проявляється у мікроорганізмів.
Спектр дії захисту полягає у 310-340 нм.
Механізм захисту пов'язаний з інгібуванням поділу клітин, зумовлений фотодеструкції речовин хінонної природи.