О
зони іннервації нервами: 1
— блукаючим; 2 —
язикоглотковим; 3 —
барабанною струною лицевого
нерва; 4 — сосочки
різного типу, то сприймають: + —
гірке; о — кисле;
І —солоне; ~ —солодке
Мал. 23. Розміщення сосочків та іннервація язика людини:
Смакові волокна відповідних черепних нервів закінчуються в ядрі одинокого шляху. Нейрони цього ядра зв'язані з руховим подвійним ядром довгастого мозку, а також через присередню петлю — з задньо-присереднім ядром таламуса. Звідси аксони нейронів третього порядку проходять крізь внутрішню капсулу і закінчуються в зацентральній звивині кори головного мозку, де знаходиться центр смаку.
Процеси, що відбуваються в цих структурах, значною мірою відповідають за виникнення певного емоційного стану організму, пов'язаного з тим чи іншим смаковим або нюховим подразником.
С
Мал.
23.1.
Схема
специфічного каналу передачі інформації
смакової сенсорної системи. 1
- смакові
рецептори язика, 2
- аферентні
черепні нерви VІІ,
IX,
X
пари,
3
-довгастий
мозок (ядро одиночного тракту), 4
- ядра
таламуса, 5
- сенсорна
смакова
зона кори.

Шлях передачі інформації від смакових рецепторів до центрів кори головного мозку - це специфічний канал передачі інформації. Обробка інформації здійснюється на рецепторному рівні, центрах довгастого мозку, ядрах зорових горбів, смакової сенсорної зони кори головного мозку (задня центральна закрутка). Участь сенсорних і асоціативних зон кори забезпечує формування смакового відчуття (мал. 23.1).
Смакові відчуття. У 1916 р. німецький вчений Хеннінг запропонував концепцію чотирьох основних смакових відчуттів: солоного, кислого, солодкого і гіркого. Проте ще у 1909 р. японець Ікеда виділив смак глутамінової кислоти (умамі), який нагадує смак курячого бульйону, а французька дослідниця Анік Форміон виділила шостий смак — гліцєризової кислоти (складова частина кореня солодки). Вона вважає, що язик людини має приблизно 10 видів рецепторів для відчуття смаку різних типів.
Вважають, що відчуття солодкого пов'язане з нуклеотидциклазою, гіркий смак виникає внаслідок виходу з рецепторних клітин внутрішньоклітинного кальцію. Відчуття кислого і солоного, які виникають під впливом малих йонів, здійснюються завдяки блокаді (кислий) чи активації (солоний) існуючих каналів на апікальній мембрані смакових рецепторів.
Багатьом природним подразникам властиві змішані відчуття, що відповідають природі їхніх компонентів. Так, помаранч має кисло-солодкий смак, а грейпфрут — кисло-солодко-гіркий. Речовинами з кислим смаком є кислоти, а з гірким — рослинні алкалоїди (хінін, нікотин). Смак гіркого відчувається основою язика, а інші смакові подразники діють на бічні поверхні язика та його верхівку (див. мал. 23).
Між хімічними властивостями речовини та її смаком немає зв'язку. Так, солодкий смак має не тільки цукор, а й солі плюмбуму (свинцю), а найсолодшими речовинами є штучні замінники цукру, які солодші за нього у десятки тисяч разів (сахарин солодший за цукрозу в 1000 разів, аспартам — у 200 000 разів).
Якість відчуття може залежати від концентрації речовини. Так, натрію хлорид у малих концентраціях видається солодким. Найбільшу чутливість язик має до гіркого (8 • 10-6 М/л), найменшу — до солодкого і солоного (1 • 10-2 М/л). Отже, завдяки високій чутливості до гіркого ми отримуємо інформацію про наявність гірких речовин у воді чи їжі, навіть якщо їх там дуже мало. А оскільки такі речовини найчастіше отруйні, то це є застереженням для живої істоти. Сильні гіркі подразники можуть навіть викликати блювотний рефлекс.
Біологічна роль смакових відчуттів полягає у контролі їстівних якостей їжі, а також у прямому впливі на процес травлення. Смакові відчуття рефлекторно пов'язані з секрецією травних залоз і зумовлюють не тільки інтенсивність секреції, а й її якість — залежно від смакових компонентів їжі. З віком здатність розрізняти смак погіршується. Кофеїн і нікотин також знижують смакову чутливість.
У різних людей абсолютний поріг смакової чутливості до різних речовин може істотно відрізнятися, аж до "смакової сліпоти" щодо окремих речовин (наприклад, фенілтіокарбамід). Крім того, абсолютний поріг смакової чутливості багато в чому залежить від стану організму (голодування, вагітність тощо). Поріг розрізнення збільшується також з підвищенням концентрації речовини. Так само люди мають різну чутливість до солодкого. Для 60% людей фруктоза є такою ж солодкою, як і цукроза, проте декому фруктоза видається солодшою у 10 разів. Розчин фруктози видається солодшим при 0°, ніж при 22°С, а також тоді, коли він змішаний з лимонним соком.
Деякі лікарські засоби можуть блокувати чи навіть змінювати смакові відчуття. Так, калію гімнемат (з індійської рослини Gimnea sylvestra) блокує сприймання солодкого (цукор викликає відчуття піску в роті). Плоди африканської рослини Synsephalium dulcificum змінюють кислий смак на солодкий (лимон сприймається як помаранч).
У разі тривалої дії смакових речовин відбувається адаптація (зниження смакового відчуття) до них. Існує також перехресна адаптація, коли зміна чутливості до однієї речовини впливає на відчуття іншої. Так, адаптація до гіркого підвищує чутливість до кислого й солоного, адаптація до солодкого поліпшує сприймання всіх інших смаків. При змішуванні кількох смакових речовин може виникнути відчуття нового смаку, який відрізняється від смаку складових компонентів.
У природних умовах існування смакові відчуття комбінуються з нюховими, тактильними, термічними, які створюються їжею. Вибір певної їжі зумовлюється не лише природженими, а й значною мірою виробленими у процесі онтогенезу умовнорефлекторними реакціями.
СОМАТОВІСЦЕРАЛЬНА СЕНСОРНА СИСТЕМА

Поняття "соматовісцеральна сенсорна система" об'єднує соматичну (тактильну, температурну, больову, пропріоцептивну) і вісцеральну (інтероцептивну) сенсорні системи. Спільним для всіх цих систем є те, що рецептори, які забезпечують цю чутливість, не створюють окремих органів чуття, а розміщені по всій поверхні і всередині тіла.
Соматична сенсорна система зосереджена у покривах тіла, починаючи з мембрани одноклітинних організмів. Через зовнішній покрив і його похідні отримується інформація про фізичні властивості навколишнього середовища. Покриви тіла виконують різні функції: захисну, видільну, обмінну,опорну, дихальну, а також сенсорну.
СОМАТИЧНА РЕЦЕПЦІЯ
Є
Мал. 24. Будова і розміщення
механорецепторів у шкірі, не вкритій
волоссям (а),
і волосистій (б):
А — надшкір'я,
або епідерміс (верхній шар — роговий);
Б —
шкіра; В— підшкірна
основа (клітковина); 1
— дотикове некапсульоване тільце
(Мейсснера); 2 —
дотиковий меніск (диск Меркеля); 3
— пластинчасте тільце
(тільце Фатера-Пачіні); 4
-рецептор волосяного
фолікула; 5
— дотиковий диск, (клітина Меркеля); 6
— дотикове капсульоване нервове тільце
(тільце Руффіні)
МЕХАНОРЕЦЕПЦІЯ
У шкірі й підшкірній основі (клітковині) знаходяться спеціалізовані нервові закінчення, чутливі до дотику, тиску, а також до механічних коливань у діапазоні 6 — 2000 Гц. У людини рецепторна поверхня шкірного аналізатора величезна — від 1,4 до 2,1 м2. У шкірі з волосяним покривом (90% усієї поверхні) основним типом рецепторів вважають дотикові (тактильні) рецептори у вигляді вільних нервових закінчень уздовж дрібних судин чи волосяного фолікула. Ці рецептори забезпечують високу дотикову чутливість.
У решті шкірного покриву (10%) — у дермі пальців кінцівок, на долонях, підошвах, губах, язиці, статевих органах і сосках грудей розміщені некапсульовані дотикові нервові тільця (Мейсснера), а також глибше розміщені пластинчасті тільця (Фатера-Пачіні) (мал. 24) — рецептори тиску й вібрації, які мають у людини розмір 0,5 — 0,7 мм.
Узагальненою моделлю механочутливого органа можна вважати пластинчасте тільце (Фатера-Пачіні), на прикладі якого розглянемо механізм механорецепції. Своєю будовою воно нагадує цибулину (мал. 25, а) і складається з 7 шарів зовнішньої цибулини, внутрішньої цибулини та аферентного немієлінізованого нервового закінчення (терміналі), яке входить у цибулину. Уздовж нервового закінчення, у пальцеподібних виростах 0,3 — 3,0 мкм завдовжки, які знаходяться в перетяжках Ранв'є, відбувається перетворення механічної енергії на електричну. Механічний стимул викликає деформацію мембрани цих виростів, її опір знижується, збільшується проникність до йонів натрію, з'являється йонний струм, далі — генерація РП (див. мал. 25, б). Після досягнення критичного рівня цього потенціалу в аферентному волокні генерується ПД, що поширюється далі.
Пластинчасті тільця належать до групи рецепторів, що швидко адаптуються. У зв'язку з цим вони адекватно реагують лише на коливання механічного подразнення частотою 30 — 200 Гц, тобто сприймають вібрацію. Механорецептори з повільною адаптацією (дотикові меніски — диски Меркеля, капсульовані дотикові тільця — тільця Руффіні) реагують на постійні подразнення — вони є рецепторами тиску. Проміжну групу утворюють рецептори з помірною швидкістю адаптації — дотикові закінчення волосяного фолікула і некапсульовані дотикові тільця (Мейсснера). Вони реагують на подразнення, що рухається, і тим активніше, чим швидкість руху більша.
Д
Мал. 25. Будова
пластинчастого тільця (Фатера-Пачіні)
(а) і
його електричні реакції на механічне
подразнення (б):
І — без
поляризації струмом, II
— з поляризацією анодним
струмом; 1, 2, 3 —
вузли нерпового волокна
(перетяжки Ранв'є); 4 — рецепторний
потенціал; N1,
N2r
N3
N1+2
— фази реакцій, які виникають
у відповідних вузлах нервового волокна
ТЕРМОРЕЦЕПЦІЯ
Температурні рецептори шкіри відіграють важливу роль у процесах пристосування організму до перепадів температури зовнішнього середовища. Вони викликають переважно виникнення місцевих судинорухових реакцій у шкірі, зумовлюють сприймання температури середовища і створення температурного відчуття. Участь у регуляції температури тіла беруть лише холодові рецептори, які в разі тривалого і значного охолодження шкіри можуть через терморегуляторний центр посилювати процеси теплотворення в організмі.
Механізми терморецепції. Терморецепторами є, ймовірно, вільні нервові закінчення, тобто вони є первинночутливими рецепторами. У багатьох випадках вони виявляють чутливість і до механічних, хімічних, больових подразнень. За спеціалізацією сприймання переважно термічної інформації терморецептори поділяються за температурним діапазоном на холодові й теплові. Холодові рецептори розміщені у поверхневих шарах шкіри і їх значно більше, ніж теплових, розміщених глибше. Крім шкіри терморецептори розміщені також у внутрішніх органах; вони сприймають температуру "ядра" тіла.
Під час реєстрації ПД у нервових волокнах від окремих терморецепторів встановлено, що всі вони тонічно активні й за сталої температури генерують ПД сталої частоти. Теплові й холодові терморецептори функціонують у різних температурних діапазонах: теплові — в діапазоні 25 — 47°С з максимальною частотою ПД при 38 — 43°С, холодові — в діапазоні 10 —40°С з найбільшою частотою імпульсів при 27—34°С.
Механізм терморецепції ґрунтується на залежності роботи йонних насосів від температури. Нагрівання холодових терморецепторів підвищує активність натрієвих насосів так, що при 30°С і вище виведення Na+ з нервового волокна спричинює його гіперполяризацію і гальмування спонтанної активності. При температурі до 25°С насос припиняє працювати і збудливість терморецептора визначається натрієвими дифузійними струмами. Теплові рецептори, ймовірно, працюють за таким самим принципом, але з калієвим насосом.
Розрізняльний поріг терморецепторів становить 0,2°С, тобто терморецептор реагує зміною електричної активності внаслідок зміни температури на 0,2°С. Поріг відчуття зміни температури залежить також від швидкості її зміни. Коли швидкість зміни температури перевищує 0,1°С за 1 с, це реєструється рецепторами. Проте при зменшенні цієї швидкості відбувається досить різке збільшення порога відчуття як тепла, так і холоду. Так, при охолодженні шкіри зі швидкістю 0,007°С за 1 с від початкової точки 33,5°С відчуття холоду виникає, коли температура ділянки шкіри знижується на 4,4°С.
Центр терморегуляції. Інформація від терморецепторів шкіри і внутрішніх органів про температуру поверхні тіла чи його ядра через бічні спинномозково-таламічиі провідні шляхи надходить до гіпоталамуса, де розміщений центр терморегуляції, і до соматосенсорної зони кори великого мозку, де формуються температурні відчуггя. В гіпоталамусі йде обробка цієї інформації, проте головну роль у формуванні команд на запуск реакцій теплотворення чи тепловіддачі відіграють центральні терморецептори — нейрони присереднього передзорового ядра гіпоталамуса, безпосередньо сприймаючи температуру цереброспіпальної рідини, що їх омиває. їх роль у підтриманні сталої температури тіла у 8—10 разів більша, ніж периферичних терморецепторів. Центральні терморецептори так само, як і периферичні, поділяються на теплові й холодові.
Температурне відчуття. Розрізняють два види температурного відчуття: динамічне (зміна температури) і статичне (сталий рівень температури). Коли людина потрапляє у середовище з температурою 22 — 24°С, то відчуття тепла з'являється тільки тимчасово, а потім ні саме тепло, ні помірні зміни температури вона не відчуває. Цю температурну зону називають зоною температурного комфорту. Для оголеної людини зона комфорту є досить вузькою — 33 — 35°С.
Адаптація до температур поза зоною комфорту відбувається повільно (десятки хвилин) і ніколи не буває повною. Саме завдяки такій неповній адаптації обстежувані особи здатні досить точно оцінювати довколишню температуру.
Температурні відчуття пов'язані зі складними процесами у ЦНС, насамперед з явищами сумації збудження. При цьому враховується активність як холодових, так і теплових рецепторів. На користь твердження, що відчуття тепла чи холоду "обчислюється" в центрі терморегуляції, свідчить те, що адаптація до нової температури на рівні суб'єктивних відчуттів займає десятки хвилин, а на рівні терморецепторів — кілька секунд.
У людини існує особлива температурна чутливість у вигляді слідового відчуття. Так, якщо притиснути на 20 — 30 с до лоба металеву лінійку, а потім забрати її, то суб'єктивне відчуття холоду зберігається протягом десятків секунд, хоча об'єктивні вимірювання свідчать про те, що шкіра вже давно нагрілася. Слідове відчуття холоду пов'язане з тривалою активністю холодових рецепторів.
Надмірні теплові стимули можуть викликати парадоксальне відчуття холоду. Це також пояснюється особливостями холодових рецепторів, які в нормі припиняють функціонувати при 40°С, але під час нагрівання до 43°С і вище короткочасно збільшують частоту розрядів ПД. Цим пояснюється виникнення відчуття холоду в разі швидкого занурення у гарячу воду.
Хоча адекватним подразником для терморецепторів є абсолютне значення температури, проте відчуття зміни температури залежить від початкової температури шкіри, швидкості й напрямку зміни температури середовища і площі шкіри, на яку діє температура. Залежно від умов стимуляції та сама температура може суб'єктивно відчуватися як тепло або холод. Так, якщо початкова температура становить 32°С, то нагрівання на 0,5°С викликає відчуття тепла, а якщо 33°С, то охолодження на 0,5°С зумовить чітке відчуття холоду, хоча кінцева температура в обох випадках є однаковою — 32,5°С.
БОЛЬОВА РЕЦЕПЦІЯ
Біль є суб'єктивним відчуттям людини. Біль є дуже специфічною сенсорною модальністю, що спричинює особливий психофізіологічний, мотиваційно-емоційний стан людини. На відміну від усіх інших модальностей біль надає організму дуже бідну, але надзвичайно важливу інформацію. Вона сигналізує організму про небезпеку ушкодження, руйнування і вмикає ланцюг захисних реакцій, що є елементами програми больової поведінки.
Біль відрізняється від інших відчуттів тим, що зумовлює негативний емоційний стан і через різні рівні ЦНС формує багатокомпонентні реакції організму. Наприклад, під час сильного болю під впливом імпульсації від больових рецепторів підвищується тонус симпатичної нервової системи, внаслідок чого через α-адренорецептори скорочується м'яз — розширювач зіниці й зіниці розширюються.
Щ
Мал. 26. Категорії
болю
Класифікація больових відчуттів. За низкою ознак біль поділяють на соматичний і вісцеральний (мал. 26). Коли соматичний біль виникає на поверхні шкіри, його називають поверхневим, а якщо від м'язів, кісток, суглобів — глибоким. Прикладом глибокого соматичного болю є головний біль. У літературі описано понад 20 видів головного болю, одним з яких є мігрень. Вона періодично повторюється, звичайно є однобічною, має чіткий початок і кінець, причина її невідома.
Якщо вколоти шкіру голкою, тобто викликати поверхневий біль, то спочатку відчувається ранній біль, який легко локалізується, а після цього, особливо при значних стимулах, виникає пізній біль з латентним періодом 0,5 — 1 с. Це тупий (ниючий) тривалий біль, який досить важко локалізувати.
Вісцеральний біль, що охоплює внутрішні органи, також буває тупим чи дифузним, він схожий на глибокий соматичний біль.
Вісцеральний біль викликається імпульсацією від больових рецепторів у внутрішніх органах. Він, по суті, є глибоким болем і здебільшого тупим, дифузним, погано локалізованим. Так, у серцевому м'язі у випадках локального порушення кровотоку (ішемія серця) виникають больові відчуття, які сприймаються людиною як загруднинний біль або біль під лівою лопаткою (стенокардія, застар. — грудна жаба). Гладком'язові стінки порожнистих внутрішніх органів при їх сильному розтяганні також спричинюють вісцеральний біль, що поширюється на всю черевну порожнину.
Больові рецептори (ноцицептори) морфологічно є первинночутливими вільними нервовими закінченнями. За характером сприймання стимулів розрізняють специфічні й полімодальні больові рецептори (термо-, механо-, механотермоноцицептори); за механізмом збудження — механо- і хемоноцицептори.
Больова чутливість шкіри є нерівномірною: больових точок з полімодальними больовими рецепторами у 8 — 10 разів більше, ніж точок, чутливих лише до тиску чи змін температури. Отже, можна говорити про існування специфічних больових рецепторів, які реагують тільки на інтенсивні стимули, викликаючи відчуття болю.
Відома виняткова больова чутливість тих тканин, де переважають вільні нервові закінчення (рогівка ока, пульпа зуба тощо). Оскільки серед вільних нервових закінчень є специфічні низькопорогові механо- і терморецептори, ймовірно, що їх сенсорну модальність визначають не будова чи форма, а певні хімічні чинники.
Механізм збудження больових рецепторів при больових впливах не з'ясовано. Припускають, що велике значення мають зміни рН тканини у ділянці нервового закінчення: біль виникає при зниженні рН, починаючи з 6 і нижче. Тому причиною виникнення болю вважають підвищення концентрації Н+ у разі токсичного чи термічного впливу на дихальні ферменти або механічного чи токсичного ушкодження клітинних мембран. Однією з причин тривалого жагучого болю може бути виділення внаслідок ушкодження клітин гістаміну, брадикініну, протеолітичних ферментів, які впливають на глобуліни міжклітинної рідини і призводять до утворення деяких пептидів, що збуджують немієлінізовані нервові волокна.
Збудження від больових рецепторів аферентними волокнами груп Аδ та С надходить до спинного мозку, далі бічним спинномозково-таламічним шляхом прямує до проміжного мозку. Відчуття раннього і пізнього поверхневого болю пов'язане з різною швидкістю проведення збудження до ЦНС. Так, у тонких мієлінізованих волокнах групи Аδ збудження поширюється зі швидкістю 15 — 30 м/с, спричиняючи раннє і точно локалізоване больове відчуття, а немієлінізовані волокна групи С зі швидкістю 0,5 — 2 м/с створюють більш пізнє, тупе відчуття болю.
Волокна групи Аδ прямують до задніх рогів спинного мозку, де переключаються на наступний нейрон, аксон якого прямує до вентробазальних ядер таламуса. Частина больових, а також тактильних волокон від цих ядер досягає соматосенсорної зони кори великого мозку, де відбувається точне визначення локалізації болючої ділянки тіла.
На відміну від цього волокна групи С у задніх рогах спинного мозку перемикаються на кілька послідовних нейронів. Медіатором, ймовірно, є субстанція Р, яка, будучи пептидом, повільно руйнується і тому довго перебуває в синаптичній щілині, що спричинює тривалу післядію. На думку багатьох дослідників, саме цією обставиною і зумовлена значна тривалість пізнього болю, який продовжує відчуватись навіть після припинення больового подразнення. Далі волокна, що проводять пізній біль, закінчуються переважно в різних структурах стовбура мозку, а також у внутрішньопластинкових ядрах таламуса. Ці структури не мають соматотонічної організації, і тому пізній біль локалізується дуже погано.
Із зазначеного випливає, що існує щонайменше два можливих механізми і відповідно дві теорії больової рецепції.
За теорією інтенсивності, рецептори будь-якої модальності можуть сигналізувати про біль за умови значної інтенсивності подразнення. При цьому автоматично знімається питання про специфічні больові рецептори, оскільки будь-який подразник достатньої сили може спричиняти біль.
Друга теорія — теорія специфічності виходить з існування специфічних больових рецепторів з відповідними провідними шляхами і центрами. Докази на користь останньої теорії було розглянуто вище.
За теорією ворітного контролю, у спинному мозку існує механізм, який постійно пригальмовує потік до ЦНС аферентних сигналів, у тому числі й больових. Це своєрідні ворота, фільтр. Больові імпульси прямують волокнами С до спинного мозку і далі до таламуса, одночасно пригнічуючи гальмівний ворітний механізм. Як наслідок відкривається широкий доступ ноцицептивних сигналів до ЦНС і больове відчуття посилюється. Теорія ворітного контролю не отримала експериментального підтвердження і її було відхилено, хоча її положення про роль задніх рогів спинного мозку у сприйманні больових сигналів зберігає своє значення.
Центри болю. Нейрохірурги під час операцій на головному мозку під місцевим знеболюванням встановили, що електричне чи механічне подразнення різних ділянок кори великого мозку спричинює різноманітні відчуття, за винятком больового. Отже, біль — це єдине відчуття, яке формується за межами кори великого мозку. У людей з патологічним процесом у таламусі будь-яке подразнення шкіри викликає сильний біль. Це дало підставу локалізувати центр болю саме у цій частині проміжного мозку. Провідну роль в аналізі ноцицептивної інформації і формуванні больових відчуттів пов'язують із задньобічним вентральним ядром таламуса, одним із головних проекційних ядер системи шкірної і м'язової чутливості. Нейрони цього ядра посилають свої аксони до першої соматосенсорної зони кори великого мозку, яка так само, як і лобова й потилична частки великого мозку, може впливати на больову чутливість організму.
Було визначено ще кілька структур, які прямо стосуються больового центру. Це протибольова (антиноцицептивна) система: центральна сіра речовина середнього мозку і велике ядро шва у ділянці моста. Функція цієї системи полягає у гальмуванні больових сигналів шляхом активації гальмівного больового комплексу, локалізованого у задніх рогах спинного мозку, який блокує больові сигнали обох (ранніх і пізніх) больових каналів ще до надходження їх до головного мозку.
Давно відомий знеболювальний ефект опію, речовини рослинного походження, і його алкалоїду морфіну. Ці речовини здійснюють знеболювання через специфічні опіатні рецептори, розміщені на мембрані нейронів антиноцицептивної системи та багатьох інших відділів ЦНС. Пізніше було знайдено ендогенні опіоїди — речовини, утворювані в організмі, які також діють на опіатні рецептори. Це низькомолекулярні пептиди лей- і метенкефалін, β-ендорфін, динорфін та інші, які через центральну сіру речовину середнього мозку або прямо через гальмівний механізм задніх рогів спинного мозку гальмують біль.
Суб'єктивно інтенсивність болю залежить не тільки від сили стимулу, а й від того, якою мірою на ньому фіксовано увагу людини. Перемикання уваги може зменшити відчуття болю (особливо у дітей), а в екстремальних ситуаціях людина може навіть не відчути болю. Шок на деякий час також знімає біль. Саме в таких випадках спрацьовує антиноцицептивна система.
Особливі види болю. В окремих випадках відчуття болю може виникати не там, звідки виходить больове подразнення.
Відбитий біль виникає при захворюваннях внутрішніх органів. Це відчуття, яке викликається больовим подразненням внутрішніх органів, локалізується не лише в цьому органі, а й у віддалених ділянках — на поверхні тіла. Такий біль проектується завжди на ті ділянки шкіри, які іннервуються від того самого сегмента спинного мозку, що й ушкоджений внутрішній орган (мал. 27). Оскільки більшість органів іннервується від кількох спинномозкових сегментів, то відбитий біль відчувається у кількох дерматомах — зонах Захар'їна-Геда (мал. 28).
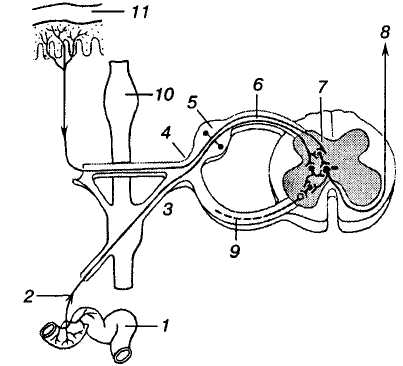
Т
Мал. 27. Рефлекторний шлях
відбитого болю: 1
— внутрішній орган; 2
— вісцеральний нерв;
3 — сполучна
гілка; 4 —
стовбур спинномозкового нерва; 5
— чутливий вузол спинномозкового
нерва; 6 — задній
(чутливий) корінець; 7
— задній ріг спинного мозку; 8
— бічний
спинномозково-таламічний шлях; 9
— передній (руховий)
корінець; 10 — симпатичний
стовбур; 11
— шкіра