

Властивості м’язового волокна. Будова і функції нервово-м’язового синапсу. Механізм утворення і передачі збудження, скорочення скелетних м’язів.
М’язи поділяються на три головні типи – скелетні, серцеві та гладенькі.
Скелетні м’язи побудовані з поперечно – посмугованої м’язової тканини. Без нервового подразнення не скорочується. Регулювання їхніх скорочень відбувається свідомо (довільно), не на довго, з великою затратою енергії.
Серцевий м’яз побудований із серцево - посмугованої м’язової тканини. Може скорочуватися автономно, не на довго, з великою затратою енергії.
Гладенькі м’язи побудовані із не посмугованої м’язової тканини. Розміщені в оболонка порожнистих органів, судинах. Скорочується мимовільно, на довго, з малою затратою енергії.
Для м’язів характерні фізіологічні властивості: збудливість, рефрактерний період, скоротливість.
Самостійна робота:
Скелетні м’язи побудовані із м’язових волокон. МПС волокна - 80-90 мв, критичний рівень поляризації біля -50 мв. ПД виникає на постсинаптичній мембрані (кінцевої пластинки) м’язового волокна, розповсюджується по мембрані у дві сторони від синапсу. Протяжність ПД в більшості 2-3 мс, швидкість розповсюдження біля 3-5 м/с. Через деякий час після ПД м’язове волокно починає скорочуватися.
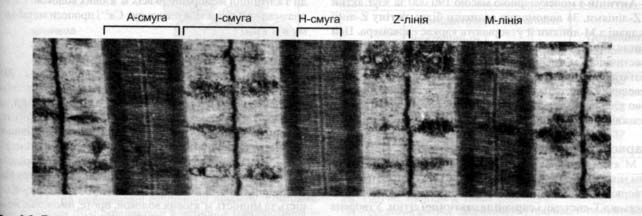
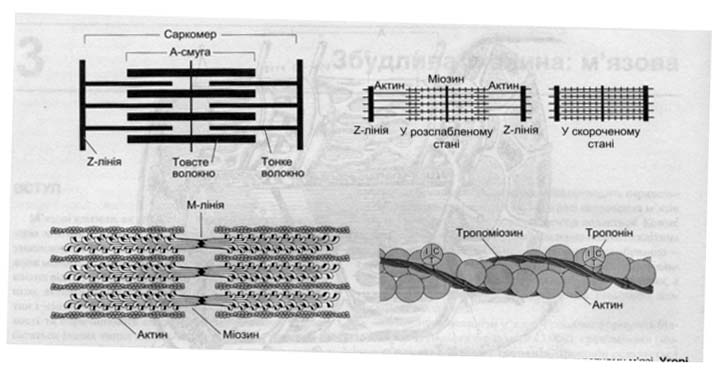
М’язове волокно в діаметрі 0,1 мм, довжина до 12 см. Під світовим мікроскопом має поперечну смугастість. Темні диски (А - анізотропні), світлі (І-ізотропні) Частина м’язового волокна від середини одного ізотропного диска до середини другого називають саркоміром.
Світлу І смугу перетинає темна Z- лінія, а темну А- смугу в центрі містить світлу Н - смугу.
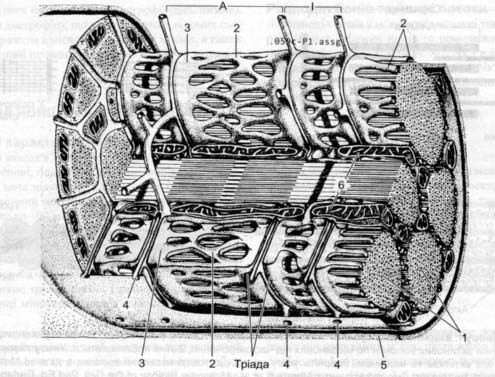
Сарколема (мембрана) має типову будову, укріпляється сполучно тканинними волокнами, які утворюють сухожилок. Саркаплазма (цитоплазма) має типовий набір органел, за винятком саркоплазматичного ретикула (СР) або називають Т системою (тріада).
СР має систему трубочок, цистерн і канальців, які оточують кожну міофібрилу СР – депо Са 2+. В саркоплазмі знаходиться білок міоглобін який є джерелом кисню.
В середині волокна розміщені скоротливі міофібрили: товсті (міозинові), тонкі (актинові).
Міозинові міофіламенти – мають головку, шийку. Головка направлена під кутом від центра в сторону активних ниток. Актинові міофіламенти побудовані із двох актинових ниток з активними центрами до яких прикріплюється головка міозина. Також міститься тропоніновий комплекс з тропоніном і тропоміозином, який закриває актиновий центр, щоб не з’єднувалась головка міозина з актиновим центром. Кожний товстий міофіламент оточений шістьма тонкими, а тонкий може з’єднуватися з трьома товстими.
Тонкі нитки переходять через смугу Z у сусідній саркомі
Механізм м’язового скорочення
ПД по мембрані по каналах Т –системи заходить у м’язове волокно і на мембрану цистерн СР. Деполяризація мембрани цистери СР відкриває електрозбудні кальцієві канали, Са 2+ починає надходити в цитоплазму. Кальцій становиться ініціатором м’язового скорочення.
Білок тропонін приєднавши Са 2+ активує АТФазу, звільняє активний центр актину від тропонінового комплексу і відбувається приєднання головки міозина до активного центру. В результаті нитки міозина підтягуються до актину; повертається міозинова головка, з використанням енергії АТФ.

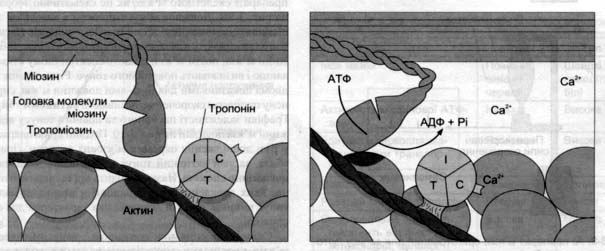
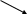
«Шарнірний механізм»
Скорочення м’язів відбувається по принципу «шарнірного механізму». З’єднання головки міозина з центром актина проходить поворот її на 45° і вона виправляється. Нове надходження Са 2+ приводить до руху міозина по актину «стрибок або крок» І так крок за кроком.
При скороченні м’язів:
а) актин і міозин не зменшується:
б) іде ковзання (або входження) одних ниток вздовж других;
в) дві сусідні Z- ліній зближаються (максимум у 2 рази);
г) при зменшенні довжини м’яза саркомір розширюється;
д) аналогічні процеси відбуваються у всіх саркомірах м’язового волокна, тому кінці м’яза підтягуються до центру.
Розслаблення м’язів
Кроки ковзання міозина і актина буде іти до тих пір поки буде достатня кількість Са 2+, АТФ. Нагромадження Са 2+ включає Са+ - насоси для роботи яких потрібно також АТФ. Коли відкачується Са 2+ і зменшуються запаси АТФ розриваються зв’язки актина з міозином відбувається розслаблення.
Нервово – м’язовий синапс закінчується присинаптичною бляшкою, яка насичена синаптичними (до 3000 000) міхурцями. Міхурці містять ацетилхолін (АХ). Синапс має синаптичну щілину та постсинаптичну мембрану (кінцеву пластину м’язового волокна).
П реси наптичская мембрана Поры в мембране Постсинаптическая мембрана
Схема нервово – м’язового синапса і механізм проведення збудження за
допомогою медіаторів.
Пресинаптична мембрана 4. Синаптична щілина
Міхурці з АХ ( ацетилхоліни) 5. Постсинаптична мембрана
Мітохондрії 6. Ядро
7. Міофібрили
ІІ. М’язове волокно
ПД надходить до пресинаптичної бляшки відкриваються кальцієві канали і кальцій надходить до бляшки (збільшення в 100 разів)
Міхурці зливаються і рухаються до пресинаптичної мембрани, зливаються з нею і секрет міхурців попадає в синаптичну щілину. Для одного ПД звільняються до 1 мл. молекул АХ. АХ досягає постсинаптичної мембрани, і взаємодіє з ацетілхоліновими рецепторами. Результат цієї роботи відкриття іонних канальців, які мають селективний фільтр 65нм. Через них можуть проходити іони Na+,К+,Са2+, але перважно лавиноподібно поступають іони натрія, які деполяризують кінцеву пластинку м’язового волокна. Канал відкритий тільки 1 мс., тому що холінестераза руйнує ацетилхолін в синаптичній щілині. Деполяризація м’язового волокна викликає ПД.
Одне подразнення викликає деполяризацію на 0,1 -015 мв. кінцевої пластини (потенціал кінцевої пластини ПКП) м’язевого волокна. Одне подразнення може не викликати ПД, а викликає ПКП.А декілька нервових імпульсів приводить до накладання величини ПКП один на одного. Їхня сумація приводить до ПД м’язового волокна. Синаптична затримка у нервово - м’язевому синапсі становить від 0,5 до 5мс.
М.Є. Введенський установив, що збільшення частоти і сили подразнення (з 10 до 50 імпульсів) приводить до збільшення амплітуди м’язових скорочень. Подальше збільшення частоти і сили подразнень приводить до зменшення амплітуди м’язових скорочень. Зменшення частоти сили подразнень приводить до відновлення м’язових скорочень.
(Закон залежності м’язових скорочень від сили і частоти нервових подразнень).
Функції скелетних м’язів:
функціонує опорно - рухова система;
забезпечує пристосованість до навколишнього середовища;
піддержується тонус центральної нервової системи;
приймає участь в теплорегуляції;
є органом відчуття;
приймає участь в водно - сольовому обміні;
прояв емоцій;
є депо глікогена, кисню.
Фізіологічні властивості:
а) збудливість менша ніж у нервової системи;
б) провідність становить 5 м/с;
в) рефрактерний період 35мс.
Існує два типи м’язових волокон тип 1, тип 2. Є червоні м’язи у яких переважають волокна типу 1, вони виглядають темнішими, повільно реагують, мають триваліший латентний період, призначені для тривалих повільних скорочень, головна функція яких підтримати стале положення тіла.
Білі м’язи побудовані переважно із волокон типу 2, вони світліші, швидко реагують, мають короткий латентний період і призначені для швидких і точних рухів.
М’язеві волокна |
Тип 1 |
Тип 2 |
Інші назви
Властивість
|
Повільні Окисні Червоні Час скорочення 100мс |
Швидкі Гліколітичні Білі Час скорочення 7,5 мс |
Активність міозинової АТФази |
Низька |
Високе |
Здатність саркоплазматичної сітки транспортувати Са2+ |
Помірна |
Висока |
Діаметр |
Помірний |
Великий |
Резерв системи гліколізу |
Помірний |
Високий |
Окислювальна здатність (вміст мітохондрій, щільність капіляр та концентрація міоглобіну) |
Висока |
Низька |
Дослідження показали що у людини є вроджена кількість швидких та повільних волокон у різних скелетних м’язів.
У стегнових м’язах кількість повільних волокон коливається від 13 –до 96 %. Звідси різниця в бігунів на короткі дистанції бігу і на довгі.
Типи скорочення: ізометричний, ізотонічний. Під час скорочення м’язів зменшується довжина скоротливих елементів, але загальна довжина може не змінюватися, таке скорочення називають ізометричним (від грецьк. Іsos – сталий,
metros – довжина.
Скорочення м’язів із наближенням їхніх кінців називають ізотонічним
(tonus – напруження).
Повторне стимулювання м’язів ПД перед їхнім розслабленням приводить до додаткового активування скоротливих елементів і в цьому разі додаткова скоротлива відповідь накладається на вже наявне скорочення. Це явище назвали
Сумацією скорочень: тонус м’язів що розвивається під час сумації, значно більший. В людини буває одне скорочення м’язового волокна, в більшості воно тетанічне.
У разі швидкого повторення подразнення активування скоротливого механізму багаторазово повторюється, перш ніж м’яз встигає розслабитися унаслідок чого окремі відповіді зливаються в одне тривале скорочення.
Такий процес називають тетанусом (тетанічне скорочення). У випадку повного суцільного тетанусу в інтервалах між подразниками не відбувається жодного розслаблення м’язів.
У разі неповного (зубчатого) тетануса скорочення під дією сумових подразників чергуються з періодами часткового розслаблення.

Електроміограма тетануса.
Совокупність рухового нейрона та м’язових волокон, які він інервує називають рухова одиниця. Кількість волокон що припадає на одну рухову одиницю різна і залежить від функції конкретного м’яза. В м’язах кисті та очного яблука тих, що забезпечують точні, високо координовані рухи, одна одиниця містить 3-10 м’язових волокон. У других м’язах можливо сотні або тисячі.
Кожен спинномозковий руховий нейрон іннервує волокно лише одного типу. Рухової одиниці поділяються на швидкі та повільні.
Повільні рухові одиниці містять невеликі за розміром нейрони із низькою швидкістю проходження імпульсу, а швидкі, відповідно, - великі нейрони із високою швидкістю проходження імпульсу.
Сила м’язів визначається силою тяжіння на її кінцях. Для вимірювання сили визначають або максимальний вантаж, який вона може підняти, або максимальний напряг, який вона може розвинути в умовах ізометричного скорочення.
Для сили м’язів характерно:
а) швидкі рухові одиниці сильніші за повільні;
б) більші за об’ємом м’язи – сильніші ;
в) косий хід волокон сильніший за прямий (більше волокон);
г) чим більший кут прикріплення до кісток тим сильніший м’яз.
Відносна сила м’язів на 1 см2 площі зрізу м’яза –
двухгологовий м’яз плеча 11,4 кг.
тригологовий м’яз плеча 16,7 кг.
Робота м’язів визначається за формулою А = m х h
де m маса , h- висота
Висота підняття залежить від маси навантаження, сили скорочення і довжини м’яза.
Робота м’язів підвищується при поступовому підвищенні маси об’єкта. Але до певного рівня, маса об’єкта і величина роботи починає знижуватися. Найбільша величина роботи відмічається при підніманні середніх вантажів.
Енергетика м’язових скорочень
Енергія м’язу необхідна для :
скорочення (утворення містків між головкою міозина і актином);
розслаблення (розрив містків);
роботи Са2 насоса;
роботи Na+,К+ насоса
Енергія використовується у вигляді АТФ. АТФ розщеплюється на АДФ і фосфорний залишок з виділенням енергії.
АТФ небагато і її хватає на декілька поодиноких скорочень. Ресинтез АТФ відбувається за рахунок: