

2. Определение катаболизма. Катаболизм и анаболизм. Стадии катаболизма биомолекул в организме.
Катаболи́зм или энергетический обмен — процесс метаболического распада, разложения на более простые вещества (дифференциация) или окисления какого-либо вещества, обычно протекающий с высвобождением энергии в виде тепла и в виде АТФ.
Анаболи́зм или пластический обмен — совокупность химических процессов, составляющих одну из сторон обмена веществ в организме, направленных на образование составных частей клеток и тканей.
В метаболизме можно выделить: пути анаболизма, которые предназначены для биосинтезов, и пути катаболизма, которые ведут к расщеплению сложных молекул. Хотя катаболические и анаболические пути во многом различаются, они тесно связаны друг с другом. Связь между ними обеспечивает оптимальный уровень метаболизма. Катаболизм и анаболизм - это сопряженные взаимодополняющие процессы.
Катаболизм сопровождается освобождением энергии, которая может аккумулироваться в виде АТФ. При анаболических процессах происходит потребление энергии, которая освобождается при распаде АТФ до АДФ и фосфорной кислоты или АМФ и пирофосфорной кислоты. Следовательно, АТФ является сопрягающим энергетическим звеном катаболизма и анаболизма. Кроме АТФ связующим звеном могут служить специфические метаболические пути или циклы. Связующий путь (цикл), объединяющий пути распада и синтеза веществ, называется амфиболическим. Примером амфиболического цикла может служить цикл Кребса. Амфиболические пути связаны, как правило, с окислением веществ до углекислого газа и воды.
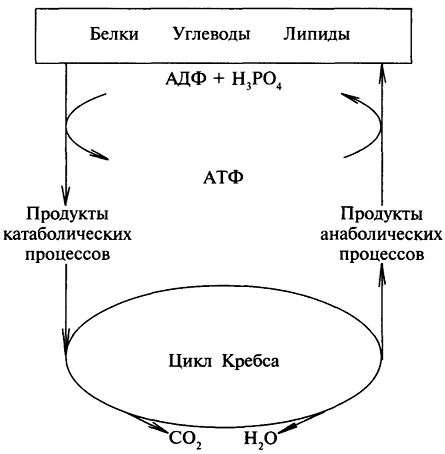
Стадии катаболизма биомолекул. При расщеплении биомолекул в организме выделяют 3 стадии, которые являются общими для катаболизма различных биомолекул.
В первойстадии все сложные биомолекулы (полимеры) расщепляются до простых компо нентов (мономеров): 1) полисахариды расщепляются до моносахаридов; 2) липиды (триа цилглицеролы) – до жирных кислот и глицерина; 3) белки – до аминокислот; 4) нуклеиновые кислоты – до мононуклеотидов. Реакции этой стадии катализируются гидролазами желудка, и кишечника. На этой стадии высвобождается около 1% химической энергии, которая рассеива ется в виде тепла.
Во второй стадии мономеры, образовавшиеся в первой стадии, внутриклеточно подвергают ся превращениям с выделением энергии (2030%). Основные реакции катаболизма:
1) для моносахаридов – гликолиз, конечным метаболитом которого является пировиноград ная кислота, которая далее подвергается окислительному декарбоксилированию и превраща ется в активную форму уксусной кислоты – ацетилКоА;
2) для жирных кислот – βокисление, конечным продуктом которого является ацетилКоА;для глицерина – расщепление до пирувата, который далее превращается в ацетилКоА;
3) для аминокислот и нуклеотидов – дезаминирование и расщепление безазотистых молекул до ди и трехуглеродных карбоновых кислот и их производных. Большинство этих метаболи тов превращается в ацетилКоА.
Таким образом, общим конечным продуктом второй стадии внутриклеточного катабо лизма углеводов, липидов и аминокислот является ацетилКоА.
В третьейстадии катаболизма в митохондриях происходит окисление ацетилКоА до СО2 и Н2О и окислительное фосфорилирование с образованием АТФ. Окисление ацетилКоА до СО2 происходит в цикле трикарбоновых кислот, при участии коферментов НАД и ФАД и ци тохромов Атомы водорода поступают в дыхательную цепь (электроннотранспортная цепь митохондрий) и переносятся на кислород, образуя Н2О. Полученная энергия (на этой стадии образуется 7080% енергии) используется для осуществления окислительного фосфорилиро вания, главного источника АТФ в организме.
3.Катаболизм глюкозы. Анаеробный и аеробный обмен. Реакции анаеробного гликолиза. Эффект Пастера. Энергетический баланс анаэробного гликолиза.
Гликолиз - это серия реакций, в результате которых глюкоза распадается на две молекулы пирувата (аэробный гликолиз) или две молекулы лактата (анаэробный гликолиз). Все десять реакций гликолиза протекают в цитозоле и характерны для всех органов и тканей. Аэробный распад глюкозы включает реакции аэробного гликолиза и последующее окисление пирувата в реакциях катаболизма.
Таким образом, аэробный распад глюкозы - это предельное ее окисление до СО2 и Н2О, а анаэробный гликолиз - это специфический путь катаболизма, т. е. часть аэробного распада глюкозы. Анаэробный распад включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата в лактат (т. е. термины анаэробный распад и анаэробный гликолиз совпадают).
В гликолизе можно выделить три основных этапа. На первом этапе превращениям подвергаются гексозы, на втором - триозы, на третьем - карбоновые кислоты.
Характеристика гликолиза:
большинство реакций обратимо, за исключением трех (реакций 1, 3, 10);все метаболиты находятся в фосфорилированной форме;источником фосфатной группы в реакциях фосфорилирования являются АТФ (реакции 1, 3) или неорганический фосфат (реакция 6);регенерация NAD+, являющаяся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. В этом случае водород транспортируется в митохондрии с помощью челночного механизма при участии переносчиков. Это происходит потому, что мембрана митоходрий непроницаема для протонов. При анаэробном гликолизе регенерации NAD+ осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от NADH является пируват, который восстанавливается в лактат;
образование АТФ при гликолизе может идти двумя путями: либо субстратным фосфорилированием, когда для фосфорилирования AДФ используется энергия макроэргической связи субстрата (реакции 7, 9), либо путем окислительного фосфорилирования AДФ, сопряженного с дыхательной цепью (реакция 6).
Анаэробный распад глюкозы (анаэробный гликолиз)
Анаэробным гликолизом называют процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому не зависит от работы митохондриальной дыхательной цепи. АТФ образуется за счёт реакций субстратного фосфорилирования. Суммарное уравнение процесса:
С6Н1206 + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 + 2 АТФ + 2 Н2O.
. Реакции анаэробного гликолиза
При анаэробном гликолизе (рис. 7-40) в цитозоле протекают все 10 реакций, идентичных аэробному гликолизу. Лишь 11-я реакция, где происходит восстановление пирувата цитозольным NADH, является специфической для анаэробного гликолиза (рис. 7-41). Восстановление пирувата в лактат катализирует лактатдегидро-геназа (реакция обратимая, и фермент назван по обратной реакции). С помощью этой реакции обеспечивается регенерация NAD+ из NADH без участия митохондриальной дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от NADH (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию NAD+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма. Это вещество выводится в кровь и утилизируется, превращаясь в печени в глюкозу, или при доступности кислорода превращается в пируват, который вступает в общий путь катаболизма, окисляясь до СО2 и Н2О. Баланс АТФ при анаэробном гликолизе
Анаэробный гликолиз по сравнению с аэробным менее эффективен. В этом процессе катаболизм 1 моль глюкозы без участия митохондриальной дыхательной цепи сопровождается синтезом 2 моль АТФ и 2 моль лактата. АТФ образуется за счёт 2 реакций субстратного фосфорилирования. Поскольку глюкоза распадается на 2 фосфотриозы, то с учётом стехиометри-ческого коэффициента, равного 2, количество моль синтезированного АТФ равно 4. Учитывая 2 моль АТФ, использованных на первом этапе гликолиза, получаем конечный энергетический эффект процесса, равный 2 моль АТФ. Таким образом, 10 цитозольньгх ферментов, катализирующих превращение глюкозы в пируват, вместе с лактатдегидрогеназой обеспечивают в анаэробном гликолизе синтез 2 моль АТФ (на 1 моль глюкозы) без участия кислорода.
Анаэробный распад глюкозы происходит в мышцах, в первые минуты мышечной работы, в эритроцитах (в которых отсутствуют митохондрии), а также в разных органах в условиях ограниченного снабжении их кислородом, в том числе в клетках опухолей. Для метаболизма клеток опухолей характерно ускорение как аэробного, так и анаэробного гликолиза. Но преимущественный анаэробный гликолиз и увеличение синтеза лактата служит показателем повышенной скорости деления клеток при недостаточной обеспеченности их системой кровеносных сосудов.
Снижение скорости потребления глюкозы и прекращение накопления лак-тата в присутствии кислорода носит название эффекта Пастера. Впервые это явление наблюдал Л. Пастер во время своих широко известных исследований роли брожения в производстве вина. В дальнейшем было показано, что эффект Пастера наблюдается также в животных и растительных тканях, где кислород тормозит анаэробный гликолиз. Значение эффекта Пастера, т.е. перехода в присутствии кислорода от анаэробного гликолиза или брожения к дыханию, состоит в переключении клетки на наиболее эффективный и экономичный путь получения энергии. В результате скорость потребления субстрата, например глюкозы, в присутствии кислорода снижается. Молекулярный механизм эффекта Пастера заключается, по-видимому, в конкуренции между системами дыхания и гликолиза (брожения) за АДФ, используемый для образования АТФ. Как известно, в аэробных условиях значительно эффективнее, чем в анаэробных, происходят удаление Piи АДФ, генерация АТФ, а также регенерирование НАД+, окисленного из восстановленного НАДН. Иными словами, уменьшение в присутствии кислорода количества Рi и АДФ и соответствующее увеличение количества АТФ ведут к подавлению анаэробного гликолиза.
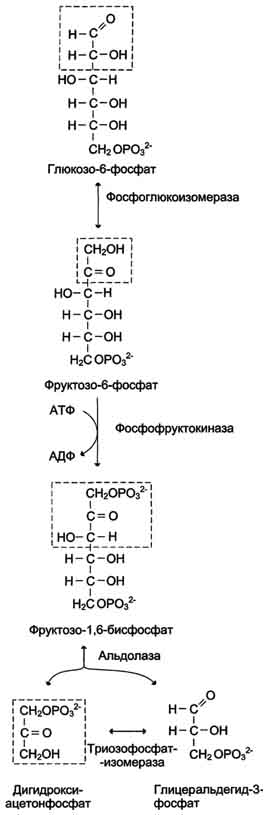
Реакции анаэробного гликолиза