

30. Особенности уф спектроскопии биополимеров.
Одна из особенностей спектроскопии биополимеров состоит в необходимости работать с растворами, поскольку изучение таких больших молекул в газовой фазе невозможно. Спектральные исследования в твердой фазе осложняются дихроичными эффектами. Для подавляющего большинства белков и нуклеиновых кислот наиболее пригодными для имитации условий in vivo растворителями являются водные буферные растворы при рН = 7,0, содержащие достаточные количества электролитов (например, ~ 0,15 М NaCl). Использование воды в качестве растворителя автоматически ограничивает спектральные измерения областью длин волн, больших чем 170 нм. Ниже этого предельного значения поглощение даже очень тонких (~ 1 мкм) водных пленок столь велико, что регистрация на его фоне вклада от макромолекул практически невозможна. Из-за высокой полярности воды электронные полосы поглощения оказываются заметно шире, чем для большинства других растворителей.
Другая трудность связана с узостью температурного интервала, в котором белки и нуклеиновые кислоты сохраняются в нативном виде, т.е. не денатурируют. Кроме того, и сама вода существует в жидком состоянии в достаточно ограниченном диапазоне 0 – 100oС. Для белков и нуклеиновых кислот это не столь важно (денатурация наступает в еще более узком диапазоне), но для модельных соединений и экспериментов невозможность проводить измерения ниже 0oС и выше 100oС оказывается подчас весьма нежелательным обстоятельством.
31. Классы хромофоров биологических полимеров.
Хромофоры белковых молекул удобно разделить на три класса - это:
пептидные группы,
боковые группы аминокислотных остатков,
простетические группы.
Для изучения свойств пептидных хромофоров можно использовать модельные соединения, такие, как формамид или N -метил ацетамид:
К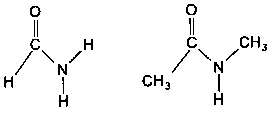 ак
известно, электроны пептидной группы
в известной мере делокализованы по трем
атомам: азоту, углероду и кислороду.
Среди наблюдаемых в пептидной группе
переходов наименьшую энергию имеет n
→ π*-переход. Как и в случае
формальдегида, n-электрон локализован
на атоме кислорода, и этот переход
запрещен по симметрии.
ак
известно, электроны пептидной группы
в известной мере делокализованы по трем
атомам: азоту, углероду и кислороду.
Среди наблюдаемых в пептидной группе
переходов наименьшую энергию имеет n
→ π*-переход. Как и в случае
формальдегида, n-электрон локализован
на атоме кислорода, и этот переход
запрещен по симметрии.
Хромофорные белки. Многие белки содержат группы, отличающиеся от обычных аминокислот. Часто, но не всегда, эти группы связаны ковалентными связями с полипептидной цепью. Вот некоторые примеры присоединенных групп: гликопротеины (сахара), гемпротеины (железосодержащие порфирины), флавопротеины (флавины), родопсин (ретиналь; витамин А). В последних трех случаях группа является хромоформом и вносит вклад в спектр поглощения в видимой или ближней УФ области.
Родопсин широко распространен в природе в позвоночных и беспозвоночных животных. Родопсин состоит из бесцветного белка (или гликопротеина), опсина, к которому ковалентно присоединена хромофорная оболочка (ретиналь). У млекопитающих изомер 11-цис-ретиналя присоединяется к ε-аминогруппе лизиновой боковой цепи опсина при помощи протонированного основания Шиффа:
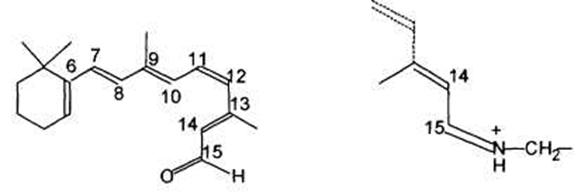
11-цис-ретиналь Основание Шиффа
Ретиналь — это пример линейных полиенов, которые мы рассмотрели ранее. Шесть сопряженных двойных связей альдегида содержат π-электроны. В результате сопряжения здесь наблюдается значительный батохромный сдвиг π → π* - перехода, что и обуславливает окраску этого хромофора.
Излучение могут поглощать содержащиеся в молекулах некоторых белков простетические группы (см. рис. 2.3.6). Так, например, гемы, флавины, пиридоксальфосфат и некоторые металлопротеиды обладают интенсивными полосами поглощения в ближней УФ- и видимой областях. По этим полосам, обычно весьма чувствительным к ближайшему окружению простетических групп, можно следить за их состоянием, например за окислением или оксигенацией. Такие хромофоры являются весьма полезными при исследовании многих ферментативных