

Види хімічних синапсів
На сучасному етапі вивчення синапсів все ще не стало можливим вироблення загальної класифікації цих структур на структурно-функціональному підґрунті. Анатомічно синапси можуть знаходитись на дендритних стовбурах, на кінцевих дендритних відгалуженнях, на тілі нейрону, або навіть на аксональних терміналах інших синапсів — в такому випадку до процесу передачі сигналу залучаються рецептори і на пресинаптичній мембрані іншого синапсу. Чітка локалізація гальмівних та збуджувальних синапсів на різних частинах нейрону простежується лише в деяких випадках. Наприклад, в спинному мозку пірамідальні нейрони утворюють інгібіторні (гальмівні) синапси на тілі клітини, а збуджувальні — на кінцевих відгалудженнях дендритів. Аналогічне анатомічне впорядкування характерне для клітин Пуркін'є в мозочку. Але, загалом, всеоохоплюючих закономірностей розташування синапсів на нейроні наразі не сформульовано.
Завдяки забарвленню іонами важких металів синапси були поділені на два типи, що часто називаються Gray-1 та Gray-2. Для типу 1 характерна наявність інтенсивного постсинаптичного ущільнення і синаптична щілина шириною 30 нанометрів; ущільнення розповсюджене практично на всю постсинаптичну мембрану (див. мікрофотографію). Тип 2 демонструє набагато менш виражене забарвлене ущільнення, і ширину синаптичної щілини, що дорівнює 20 нанометрам. Як показано в дослідах на гіпокампі та головному мозку, локалізовані там синапси типу 1 є збуджувальними, а типу 2 — гальмівними. За деякими даними, ця кореляція спостерігається також в корі та в інших частинах головного мозку ссавців, але в спинному мозку майже не спостерігається.
Надалі було показано, що початкова альдегідна фіксація препаратів мозку надає (з невідомих досі причин) синаптичним везикулам сплюснутої форми, але тільки в синапсах типу 2; радіолігандне мічення показало, що в таких синапсах із сплюснутими везикулами в якості медіатора (а, отже, вмісту везикул) використовується гамма-аміномасляна кислота (ГАМК) або гліцин — тобто інгібіторні (гальмівні) нейромедіатори. Таким чином було встановлено додатковий структурно-функціональний зв'язок для частини синапсів.
Окрім того, синапси можуть бути класифікованини за медіатором, який вони використовують ГАМК, ацетилхолін, норадреналін, опіати і т. ін.
Головні особливості хімічних синапсів
Синтез нейромедіатора у пресинаптичному нервовому закінченні.
Накопичення нейромедіатора в секреторних пухирцях.
Регульоване вивільнення нейромедіатора в синаптичну щілину.
Наявність специфічних рецепторів для нейромедіатора на постсинаптичній мембрані, які зумовлюють відповідний ефект при дії на них нейромедіатора. Аплікація нейромедіатора на синапс імітує наслідки подразнення нерва.
Наявність механізмів, що припиняють дію вивільненого нейромедіатора.
Електричний синапс
Рзповсюдження
В першій половині ХХ сторіччя вважалося, що передача нервового сигналу за допомогою хімічного синапсу притаманна периферійним нервово-м’язовим контактам хребетних (автономна мускулатура та моторні контакти), але, в основному через брак дослідних даних, синапси в Центральній нервовій системі (ЦНС) вважались такими, що використовують безпосередньо електричну передачу. Але, після відпрацювання методик внутрішньоклітинного запису електричних потенціалів в 1950-х роках, було показано, що з’єднання нейронів хімічними синапсами є правилом для ЦНС хребетних.
Надалі деякі синапси в нервовій системі ракоподібних були ідентифіковані як електричні. На теперішній час виявлена невелика кількість електричних синапсів в мозку ссавців, і велика їх доля в ЦНС нижчих хребетних, особливо риб. У безхребетних електричні синапси досить звичайні в нервовій системі аннелід та ракоподібних. Електричні синапси звичайно розташовані на таких нервових шляхах, де потрібне додержання точно визначеного часу проходження нервового імпульсу – з огляду на те, що електричний синапс не спричинює помітної в досліді затримки сигналу.
Будова
На відміну від хімічного синапсу, синаптична щілина в електричному синапсі є надзвичайно вузькою (близько 3.5 нанометри). Через синаптичну щілину даного типу синапсів проходять просторво впорядковані гідрофільні протеїнові тунелі, кожний приблизно 5 нанометрів завширшки, котрі перфорують пре- та постсинаптичну мембрану і називаються коннексонами. У первинноротих організмів (нематоди, молюски, членистоногі) коннексони сформовані протеїнами специфічної структури, що називаються паннексинами або іннексинами; у вторинноротих (голкошкірі, асцидії, хребетні) коннексони побудовані з протеїнів іншого типу – коннексинів, котрі кодуються іншою групою генів.
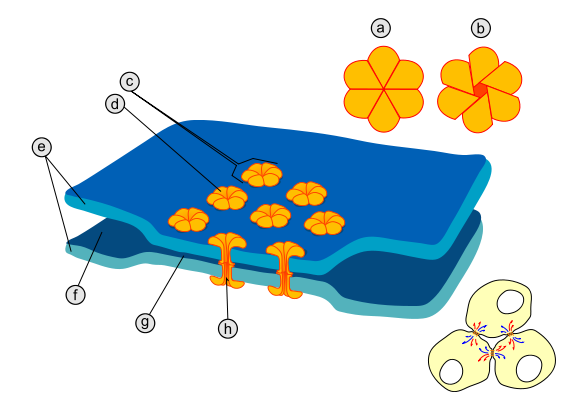
Рис.2 Будова електричного синапсу: а - коннексон у закритому стані; b - коннексон у відкритому стані; с - коннексон, вбудований в мембрану; d - мономер коннексину; е - плазматична мембрана; f - міжклітинний простір; g - проміжок у 2-4 нанометри в електричному синапсі; h - гідрофільний канал коннексону.
Ссавці в цьому плані є менш диференційовані вториннороті, і в їхньому організмі поряд з коннексинами виробляються також і паннексини, але досі у хребетних не виявлено жодного електричного синапсу, де міжклітинні канали були б сформовані паннексинами.
Міжмембранні тунелі, сформовані коннексинами (або паннексинами) забезпечують рідинний взаємозв’язок між двома нейронами – пре- та постсинаптичним - та уможливлюють прохід через них іонів та малих молекул, в тому числі штучно введених в клітину флуоресцентних барвників. Прохід вказаних барвників через електричний синапс може бути зареєстрований навіть за допомогою світлового мікроскопу.
Електричні синапси дозволяють здійснення електричної провідності в обох напрямках (на відміну від хімічних); втім, останнім часом у деяких ракоподібних було відкрито спрямовуючі електричні синапси, тобто такі, що дозволяють проходження нервового сигналу тільки в одному напрямку.