

8.5.3.2."Судьба" эмбриональных клеток и дифференцировка нейронов. Генетические мозаики и химеры. Различная "судь-
ба", т.е. направление будущей дифференцировки эмбриональных клеток-предшественниц ЦНС детерминируется на ранних этапах эмбриогенеза – на стадии бластулы у дрозофилы и стадии нервного гребня у мыши. Прослеживая судьбу поколений клеток в процессе развития ЦНСмлекопитающих, и в частности мыши, исследователи делают заключение о том, что генетически детерминированная судьба клетки сказывается не столько на окончательном положении в мозге, которое она займет как потомок клетки-родоначальницы, сколько на функциональных свойствах и количестве нейронов данного класса.
Генетические исследования развития в их традиционном варианте возможны лишь при наличии генетической изменчивости по тому или иному признаку, т.е. при выявлении и анализе эффектов соответствующих мутаций. Примером могут служить упоминавшиеся выше неврологические мутации дрозофилы – shaker, hyperkinetic (вызывающие судороги при впадении в эфирный наркоз), мутации cacophony, fruitless, stick (с нарушениями процесса спаривания), а также неврологические мутации мыши – reeler, weaver, quaking, waltzer и др.
Однако в случае очень многих мутаций экспрессия аномального гена вызывает гибель зародыша. В то же время иногда оказывается возможным реконструировать функцию нормального аллеля данного гена. Если бы можно было выделить мутации нескольких генов, экспрессирующихся на последовательных стадиях пренатального онтогенеза, это дало бы достаточно полную картину развития нервной системы. Наиболее вероятно, что такую информацию можно будет получить, изучая развитие дрозофилы, не только потому, что у нее выявлены нейрогенные и антинейрогенные локусы (см.: 8.5.3.1), но и потому, что у этого вида можно использовать чрезвычайно плодотворный метод генетических мозаиков.
Сходный методический подход у мышей обеспечил создание химерных и трансгенных животных.
Генетические мозаики. С помощью этого метода удается проследить судьбу измененных мутацией клеток и тканей на всем протяжении эмбриогенеза.
 Насекомые.
Истинными генетическими мозаиками
считаются животные, клетки которых
происходят из одной зиготы, но несут,
тем не менее, определенные генетические
различия. Наиболее известным примером
таких мозаиков являются гинандроморфные
особи, описанные у паразитических ос
Habrabracon
juglandi П.
Уайтингом. Гинандромор-фы, т.е. особи,
имеющие ткани и самца, и самки, очень
редко (1 случай из 1000– 10000) выводятся из
аномальных яиц с двумя ядрами, из которых
бывает оплодотворено только одно.
Женские ткани развиваются из диплоидного
Насекомые.
Истинными генетическими мозаиками
считаются животные, клетки которых
происходят из одной зиготы, но несут,
тем не менее, определенные генетические
различия. Наиболее известным примером
таких мозаиков являются гинандроморфные
особи, описанные у паразитических ос
Habrabracon
juglandi П.
Уайтингом. Гинандромор-фы, т.е. особи,
имеющие ткани и самца, и самки, очень
редко (1 случай из 1000– 10000) выводятся из
аномальных яиц с двумя ядрами, из которых
бывает оплодотворено только одно.
Женские ткани развиваются из диплоидного
ядра, а мужские – из неоплодотворенного, гаплоидного. Если ткани, развившиеся из оплодотворенного яйца, имеют четкие морфологические отличия, например измененный цвет глаз или аномальное жилкование крыльев, то легко видеть, что тело гинандроморфной особи представляет собой мозаику из тканей самца и тканей самки (рис. 8.18). Наблюдая за поведением таких ос, Уайтинг показал, что насекомые ведут себя либо как самец, либо как самка в зависимости от того, как распределяются на теле участки "самцовых" и "самочьих" тканей (см.: Эрман, Парсонс, 1984).
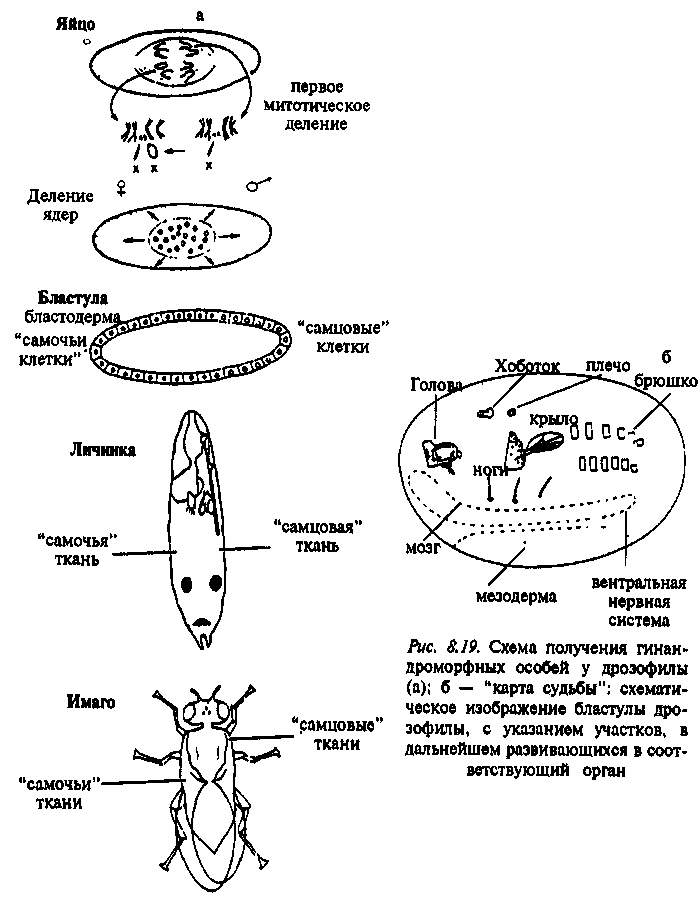
У дрозофилы мозаичные по тканям самца и самки особи получаются, когда в скрещивание берутся самки с так называемой кольцевой ^-хромосомой. Эта кольцевая структура неустойчива и часто утрачивается в период ранних митотических делений эмбрионов женского пола, которые получили одну кольцевую, а одну нормальную А1 хромосомы. Клетка, в которой остается только одна JT-xpo-мосома, дифференцируется по мужскому типу (поскольку у дрозофилы особи с генотипом ХО – самцы). Мозаицизм обнаруживается по рецессивным генам, локализованным в Х-хромосоме, а также по тем признакам, развитие которых определяется генетической программой, свойственной одному или другому полу (рис. 8.19; 8.20).
Благодаря работам С. Бензера и его коллег, проведённым в 60–70-е годы, в настоящее время имеется достаточно полная картина так называемой "карты судьбы" ("fate map") – топографического расположения на бластуле участков, из которых впоследствии развивается тот или иной орган или часть тела дрозофилы (см.: Бензер, 1975). Основные успехи ней-рогенетики и генетики поведения дрозофилы базируются именно на использовании метода гинандроморфных мозаиков.
Млекопитающие. Единственный случай сходного типа мозаицизма у позвоночных описан еще в начале 70-х годов. Это исследование было посвящено моза-ичности в проявлении мутации sh-1 мыши, затрагивающей строение кортиева органа. Анализировали фенотип самок мышей, гомозиготных по sh-1 и имеющих в гетерозиготном состоянии так называемую транслокацию Катганаха (транслокация – это хромосомная перестройка, при которой фрагмент одной хромосомы соединяется с другой). При упомянутой транслокации фрагмент 7-й хромосомы с мутантным аллелем sh-1 оказался соединенным с А"-хромо-сомой. Как известно, в каждой из клеток тела самки в активном состоянии находится только одна Af-хромосома, а вторая инактиви-рована. Поскольку в случае данной транслокации ген sh-1 был соединен с одной из А'-хромосом, а мутация эта рецессивна, то в тех клетках, где была инактивирована А"-хромосома, несущая транслокацию (и ген sh-1), эффект мутантного гена не обнаруживался. Если же была инактивирована А-хромосома, не имевшая транслокации, то оба гена sh-1 были активны и проявлялся эффект рецессивной мутации. Фенотипически это выражалось в том, что в кортиевом органе участки мутантных и нормальных клеток перемежались.

Химеры млекопитающих. Животных с генетически неоднородным организмом можно получать и в специально спланированных эмбриологических экспериментах (Мак-Ларен, 1979) – речь идет о химерах. Получение химер млекопитающих состоит из нескольких этапов. При использовании агрегационного метода извлеченные из половых путей самки эмбрионы на стадии дробления приходят в соприкосновение друг с другом, в результате чего они сливаются в один. С помощью инъекционного метода в бластоцисту эмбриона вводятся клетки другого эмбриона или другие донорские клетки (например, клетки определенной опухоли, клетки костного мозга и т.д.). На следующем этапе химерный эмбрион переносят в матку приемной матери.
Для распознавания отдельных компонентов химерного организма необходимо, чтобы ткани разного происхождения имели четкие и простые маркеры. Таким маркером может быть особенность пигментации, наличие редких форм изозимов, антигены, хромосомные транслокации и т.п.
Получение химерных животных в настоящее время налажено во многих лабораториях. Проблема подбора адекватных маркеров (морфологических – цвет шерсти, или биохимических – изофер-менты) для анализа "происхождения" того или иного участка мозга химерного животного, как правило, бывает достаточно трудной. Для такого маркирования используются, например, межлинейные генетические различия в локусе структурного гена бета-галактозида-зы. Этот фермент обнаруживается цитохимически в крупных нейронах. Нередко используют также антигенные различия в генопро-дуктах разных аллелей гена глюкозофосфатизомеразы.
Большинство неврологических мутаций мыши (это, как правило, аномалии строения мозжечка и/или внутреннего уха) рецессивны, т.е. их эффект проявляется в гомозиготном состоянии, а гетерозиготные особи практически не отличаются от нормы. В то же время особи, гомозиготные по этим мутациям, имеют резко сниженную жизнеспособность и поэтому трудны для изучения. Получение мышей-химер, составленных из нормальных и мутантных тканей, позволяет изучать мозг, мозаичный по генетическому составу. При удачном подборе маркеров можно увидеть, что даже рядом расположенные нейроны одного класса (например, клетки Пуркинье) имеют разный генотип. Как правило, неврологический дефект у таких химерных мышей не проявляется, однако на нейроанатоми-ческом уровне можно установить, какой именно тип клеток несет первичный дефект мутации, а какие клетки страдают вследствие нарушений нормальных эпигенетических отношений с первично аномальными. Данные по мутации Lurcher, упоминавшиеся выше (см.: 8.5.1), были получены именно таким методом. Анализ особенностей поведения мышей-химер также представляет интерес для выяснения роли генотипа в определении особенностей той или иной реакции. С мутацией Lurcher, например, были проведены эксперименты, показавшие, что у гетерозигот lc/+ сильно нарушена локомоция, тогда как у мышей-химер, "составленных" из эмбрионов, гомозиготных по этой мутации, и нормальных зародышей, таких изменений не было (см. также: Goldowitz et al., 1992).
На серийных срезах мозга, например на срезах ствола и мозжечка химерных мышей, у которых нормальные и мутантные нейроны различаются визуально, можно определить общее количество нейронов в моторном ядре лицевого нерва, а также число клеток Пуркинье мозжечка с нормальным и мутантным фенотипами. Статистические оценки позволили заключить, что родоначальниками клеток Пуркинье в мозге мыши являются 8 клеток, а нейроны ядра лицевого нерва происходят от 12 клеток-предшественниц.
У мышей известны межлинейные различия по числу нейронов той или иной структуры мозга. Есть веские основания предполагать, что одной из причин этой изменчивости могут быть различия в числе клонов, формирующих данный участок мозга, т.е. в числе "клеток-родоначальниц" и/или в размере клона, т.е. в числе митотических делений исходной клетки перед началом дифференцировки.
Нейрохимические, морфологические и физиологические различия между инбредными линиями мышей, описанные к настоящему времени, несомненно, определяются значительным числом локусов. Современные методы генетического анализа, использующие множественные маркеры, а также конгенные и рекомбинантные линии (в частности, метод картирования QTL – quantitative trait loci), позволяют описывать такие межлинейные различия. Возможно, они помогут и в описании структурных различий в мозге, которые являются ключевыми в возникновении межлинейных различий в поведении.
Трансгенные мыши. На основе метода получения химерных животных был разработан принципиально новый подход к исследованию генетических закономерностей формирования организма животного. Он получил название "метода создания трансгенных животных", о котором говорилось в разделе 8.5.2. Этот метод позволял исследовать особенности развития мозга и поведения животных, у которых были искусственно изменены или "выбиты" (отсюда название "нокаут", knock-out) ген или гены, кодирующие определенные структурные и/или регуляторные белки (см.: 8.6.3.3 и 8.6.4.3).