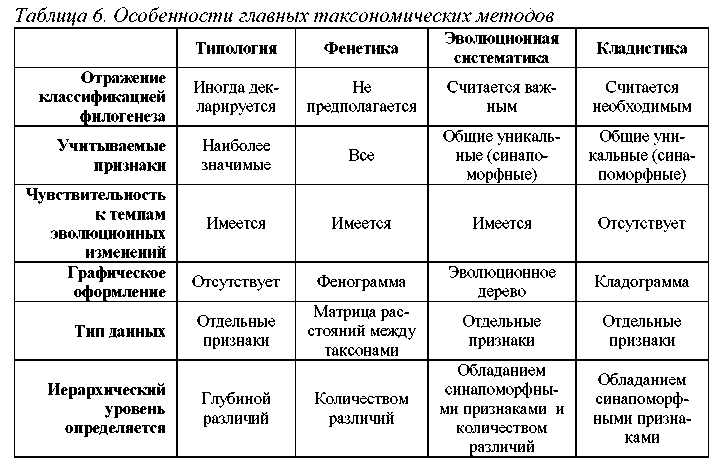
1. Должна ли классификация отражать ход филогенеза?
Типологии и фенетики не считают отражение филогенеза в классификации обязательным, эволюционные таксономисты используют филогенетические данные, однако не всегда придают им решающее значение, и, наконец, кла-дисты считают филогенез единственным основанием для классификации.
2. Должна ли неравномерность темпов эволюции влиять на классификацию?
Типологические и фенетические классификации чувствительны к неравномерности скоростей эволюции, т.к. длительный эволюционный процесс приводят к накоплению идиоадаптивных признаков, активно используемых как в типологии, так и в фенетике. Эволюционные таксономисты считают, что варьирование темпов эволюции следует учитывать в построении классификации. Кладисты напротив, стремятся исключить влияние темпов эволюции на классификацию. Таксономическое расстояние при этом определяется лишь числом точек дивергенции, разделяющих таксоны, но не зависит от числа различий, накопленных в результате единичной диверегенции.
3. Все ли признаки организмов полезны для построения классификации?
Типологии распределяют признаки по степени их значимости, и присваивают им соответствующий таксономический ранг. Фенетики полагают, что все признаки организмов являются таксономически равнозначными. Кладисты считают, что таксоны следует формировать только на основе уникальных признаков данной группы, остальные же признаки в ходе построения классификации можно не учитывать. Многие эволюционные систематики принимают кладистическую точку зрения на этот вопрос, но считают, что при установлении ранга таксона должны учитываться не только уникальные признаки, но и степень различия между группами по всему комплексу имеющихся признаков.
4. Каким должно быть графическое представление данных?
Типологический метод не предполагает графического оформления классификации. Фенетики оформляют классификации в виде фенограмм (кластерных или факторых диаграмм), кладисты – в виде кладограмм, а сторонники эволюционной систематики – в виде эволюционных деревьев. Для того, чтобы наглядно продемонстрировать сущностные различия между этими гра-
фическими конструкциями, рассмотрим их построение на конкретном примере.
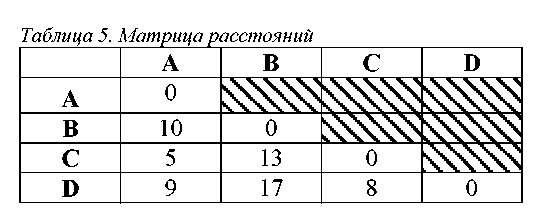
На табл. 4 представлена матрица состояний двадцати признаков у пяти анализируемых таксонов. Таксон Х является гипотетическим общим предком. Все его признаки находятся в первичном (нулевом) состоянии. В остальных таксонах признаки находятся в различных состояниях.
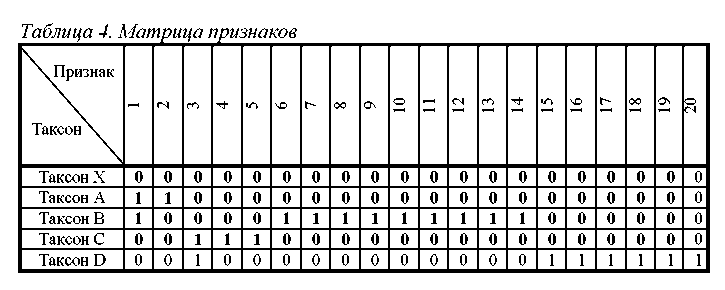
Проанализируем различия между таксонами А, В, С и D. На табл. 5 представлена матрица расстояний, описывающая степень этих различий. Приведенные в ней величины указываю по скольким признакам отличаются друг от друга таксоны данной пары. Из матрицы видно, что таксоны А и С обладают наименьшим числом различий (отличаются по 5 признакам), а таксоны В и D отличаются друг от друга в наибольшей степени (по 17 признакам). Остальные пары таксонов (А+D, А+В, В+С) занимают промежуточное положение (отличия, соответственно, по 9, 10 и 13 признакам).
Приведенные данные могут быть использованы для построения фенограммы (рис.17.а), которая объединит группы А и С, как наиболее сходные, в единый супертаксон, и противопоставит им В и D. Так будет реализован фене-тический подход к анализу рассматриваемых данных.
В отличие от фенетических, кладистические методы формируют группы только на основе общих апоморфных признаков. В рассматриваемом случае
таких признаков всего два: признак № 1 в состоянии «1», общий для таксонов А и В, и признак №3 в состоянии «1», общий для таксонов С и D. Все остальные апоморфные признаки уникальны для отдельных таксонов (например, признаки 6 – 14 у таксона В и признаки 15 – 20 у таксона D). Поэтому при кладистическом анализе они расцениваются как не имеющие отношения к процессу формирования групп. В соответствии с вышесказанным, кладограмма отношений между таксонами будет включать супертаксоны А+В и С+D (рис.17.б). Итак, кладограмма и фенограмма, построенные на основании одних и тех же данных, демонстрируют совершенно различные отношения между анализируемыми таксонами.
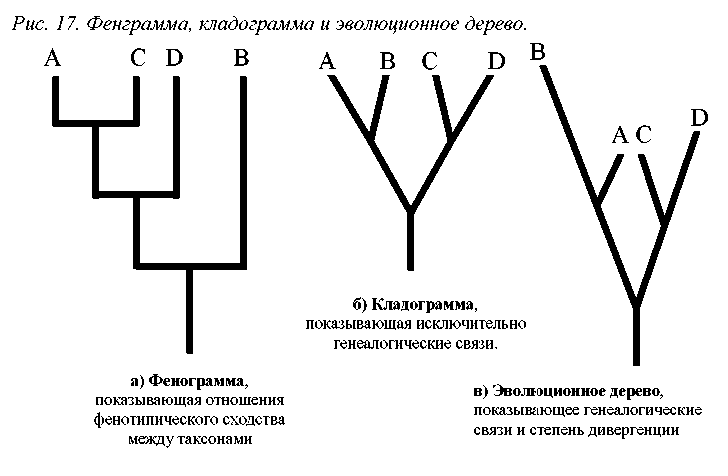
Использование метода эволюционной систематики приведет к построению дерева, близкого к кладограмме. Но согласно принципам данного направления, структура дерева должна указывать на степень и направление дивергенции таксонов. На эволюционном дереве (рис.17.в) в мы видим, что таксоны А и С, имеющие различное происхождение, сближены (так демонстрируется их фенотипическое сходство), а таксон В отстоит от основания дерева дальше, чем таксон D (так демонстрируется накопление группами различного числа уникальных признаков). Учет этих данных может вызвать своеобразную интерпретацию данного дерева, с формированием полифиле-тического таксона А+С а также парафилетических таксонов В и D, чего не в коем случае не произойдет при использовании кладограммы.
В заключение приведем таблицу, указывающую на основные особенности четырех главных таксономических методов (Табл. 6).