

Лекция 10 Мышцы и мышечное сокращение
Важнейшую роль во взаимодействии человека с внешней средой играет мышцы. Масса мышц намного больше, чем масса других органов и составляет 40-50% массы тела.
Структурные основы сокращения. Поперечнополосатые мышцы.
По морфологическим и функциональным характеристикам мышцы разделяют на два основных типа - гладкие и поперечнополосатые. Поперечнополосатые мышцы формируют двигательный аппарат скелета, глазодвигательный, жевательный и некоторые другие двигательные аппараты. Морфологически к поперечнополосатым относится и сердечная мышца. Поперечнополосатые мышцы человека и позвоночных животных (за исключением сердечной) полностью контролируются ЦНС, они лишены автоматизма. Их называют произвольной мускулатурой, имея в виду их подчинение воле человека. Гладкие мышцы позвоночных обслуживают внутренние органы. Они слабо контролируются ЦНС, обладают автоматизмом и собственной интрамуральной (внутренней) нервной сетью, в значительной степени обеспечивающей их самоуправление. Гладкую мускулатуру, а также мышцу сердца иногда называют непроизвольной.
Поперечнополосатые мышцы скелета позвоночных состоят из множества отдельных мышечных волокон, которые расположены в общем соединительно-тканном футляре и крепятся к сухожилиям, связанным со скелетом. Мышечные волокна тесно прилегают друг к другу и работают параллельно. Сила мышцы при прочих равных условиях пропорциональна числу волокон, иначе площади так называемого физиологического поперечного сечения мышцы, т.е. площади поверхности, пересекающей все действующие мышечные волокна.
Диаметр волокон поперечнополосатых мышц составляет от 5 до 100 мкм, а длина может достигать 5 сантиметров и более. Эти необычные размеры обусловлены происхождением мышечных волокон в процессе индивидуального развития. Сначала у эмбриона возникают отдельные клетки - миобласты, они сливаются в миотрубочки. Из последних формируются многоядерные, гигантские клетки - мышечные волокна (рис.).
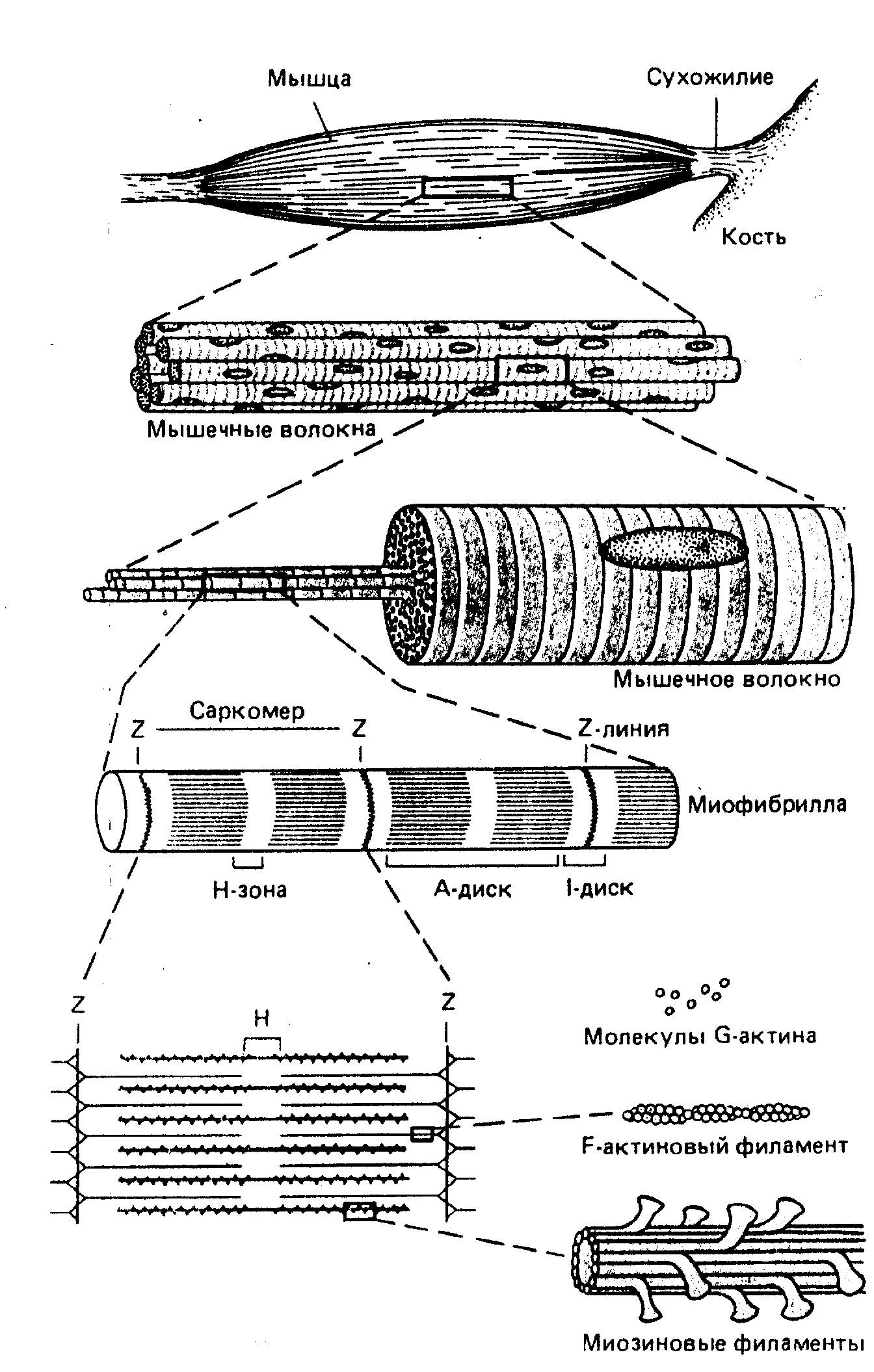
Каждое мышечное волокно заполнено множеством параллельно расположенных нитевидных образований диаметром около 1 мкм, называемых миофибриллами. В миофибриллах выделяют повторяющиеся в продольном направлении блоки - саркомеры, отделенные друг от друга Z-пластинками. Саркомер миофибрилы представляет собой функциональную, сократительную единицу поперечнополосатой мышцы, длина которого около 2,5 мкм. На электронной микрофотографии видно, что в обоих направлениях от Z-пластинки, содержащей белок α-актинин, тянутся многочисленные, до нескольких тысяч тонкие нити (филаменты), состоящие из двухнитчатой спирали F-актина, идентичного белку цитоскелета эукариотических клеток. Актиновые филаменты (диаметр 5 нм), прикреплены к Z-пластинам наподобие щетинок в щетке чередуются с толстыми нитями (диаметр 10 нм) белка миозина. Один грамм ткани скелетной мышцы содержит примерно 100 мг этих сократительных белков. Миозиновые филаменты не прикрепляются к Z-пластине и образуют наиболее плотную (анизотропную, т.е. поляризующую видимый свет) центральную часть саркомера - А-диск или А-полосу. Его длина составляет около 1,6 мкм. При тщательном электронно-микроскопическом исследовании на миозиновых филаментах обнаруживаются мелкие выступы, называемые поперечными мостиками. Более светлый участок в центре А-диска называют H-зоной. Изотропный, не поляризующий свет, светлый участок саркомера между двумя А-дисками, называют I-диском. Если сделать поперечный разрез саркомера в I-диске будут видны только актиновые филаменты (рисунок). На разрезе проходящем через h-зону видны только миозиновые филаменты. На участке взаимного перекрывания каждый миозиновый филамент окружен шестью актиновыми. В свою очередь каждый актиновый филамент окружен тремя миозиновыми. Обычно, миофибриллы отдельного мышечного волокна уложены таким образом, что расположение саркомеров на них совпадает, и на поперечном разрезе мышечного волокна у всех миофибрил видна одна и та же зона саркомера. Это создает картину "поперечнополосатости" волокна при наблюдении в световой микроскоп.
Плазматическая мембрана мышечного волокна дает на уровне границы Z-пластинки регулярные, глубокие Т-образные впячивания, представляющие собой трубки диаметром 50 нм. Эти трубки (Т-система) анатомически соединяют поверхностную мембрану с миофибриллами, лежащими в глубине мышечного волокна и обеспечивают перенос ПД с поверхностной мембраны внутрь клетки к каждой миофибрилле.
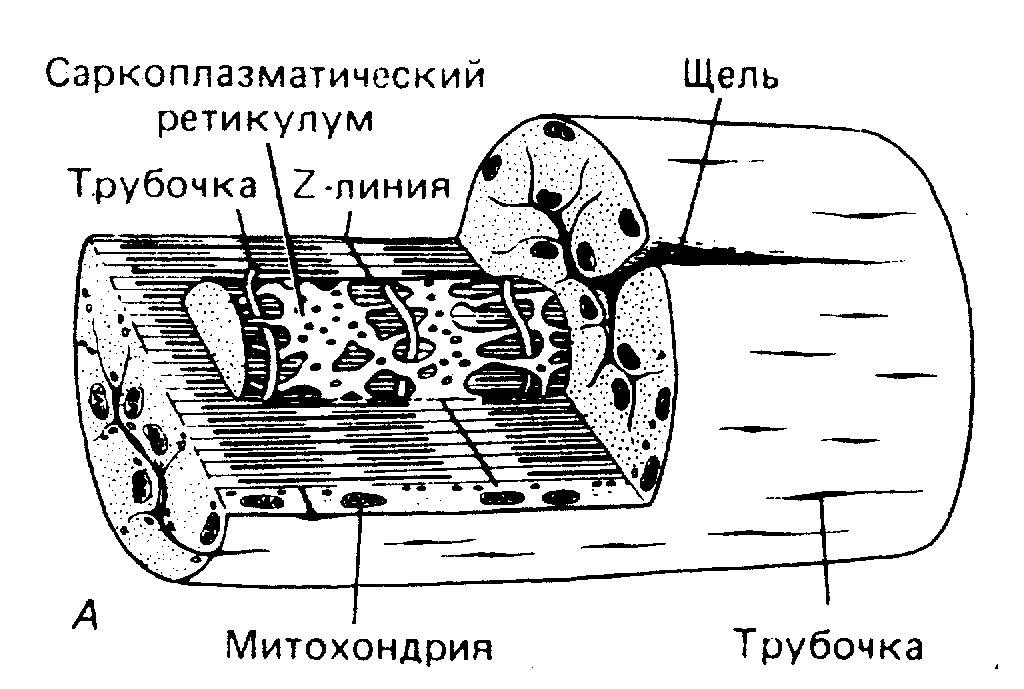
В дополнение к Т-системе в мышцах есть еще одна мембранная система, называемая саркоплазматическим ретикулумом (так называется эндоплазматический ретикулум в мышечных клетках). СР обвалакивает наподобие полой манжеты отдельно каждую миофибриллу. На уровне Z-пластины каждого саркомера цистерны СР вступают в тесный контакт с Т-трубочкой и как бы сжимают ее между собой.