

Медиаторы - точки приложения и характер действия
Одна из самых сложных задач, стоящих перед нейрофизиологами, состоит в точной химической идентификации медиаторов, действующих в различных синапсах. К настоящему времени известно довольно много соединений, которые могут выполнять роль химических посредников при межклеточной передаче нервного импульса. Однако точно идентифицировано лишь ограниченное число таких медиаторов; некоторые из будут рассмотрены ниже. Для того чтобы медиаторная функция того или иного вещества в какой-либо ткани была неопровержимо доказана, должны удовлетворяться определенные критерии:
при прямом нанесении на постсинаптическую мембрану вещество должно вызывать в постсинаптической клетке абсолютно такие же физиологические эффекты, что и при раздражении пресинаптического волокна;
должно быть доказано, что это вещество выделяется при активации пресинаптического нейрона;
действие вещества должно блокироваться теми же агентами, которые подавляют и естественное проведение сигнала.
Ацетилхолин
Наиболее известным из установленных медиаторов является ацетилхолин (уксуснокислый эфир холина).
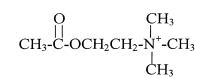
Ацетилхолин, выделяющийся окончаниями так называемых холинэргических нейронов и опосредует быструю передачу возбуждения и медленную передачу как возбуждения, так и торможения. В первом случае он воздействует на ионотропные никотиновые рецепторы или н-холинорецепторы. Свое название н-холинорецепторы получили по тому, что в экспериментальных условиях они могут активироваться никотином. Н-холинорецептор представляет собой гликопротеин, состоящий из пяти трансмембранных пептидов, имеющих участки связывания ацетилхолина. При связывании двух молекул медиатора с мультибелковым комплексом происходит конформационное изменение, приводящее к открыванию канала примерно на 1 миллисекунду. Через открытый канал способны проходить ионы, как натрия, так и калия. Так как градиент напряжения при потенциале покоя почти уравновешивает градиент ионов калия, то и сила движущая эти ионы через мембрану будет близка к нулю. Напротив, для ионов натрия, как градиент напряжения, так и градиент концентрации действуют в одном направлении, способствуя их движению внутрь клетки. Натриевый возбуждающий ток вызывает деполяризацию мембраны (ПКП) и при достижении пороговой деполяризации возникает ПД. Метаботропные рецепторы, не связанные с каналами и осуществляющие медленную тормозную или возбуждающую передачу называют мускариновыми рецепторами или м-холинорецепторами, поскольку в экспериментальных условиях они могут активироваться мускарином, одним из грибных ядов. Помимо никотина и мускарина, специфически активирующих определенные рецепторы (такие вещества называются агонистами), имеются сильнодействующие рецептор-специфические блокаторы (антагонисты), избирательно подавляющие функцию ацетилхолиновых рецепторов (ХР) того или другого типа. Например, кураре или его действующее начало D-тубокурарин и α-бунгаротоксин, белковое вещество, выделенное из яда крайта (семейство кобр), специфически связываются с н-холинорецепторами, блокируя их активность. Атропин - алкалоид красавки и белены, действует таким же образом на мускариновые рецепторы.
Из синаптической щели ацетилхолин удаляется в результате гидролиза до холина и ацетата, катализируемого ферментом ацетилхолинэстеразой (АХЭ). Этот фермент обычно находится в синаптической щели рядом с поверхностью пресинаптической мембраны (рис.).
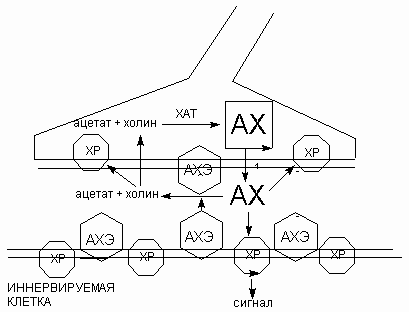
Рис. Схема функционирования холинэргического синапса
Продукты гидролиза на постсинаптическую мембрану не действуют, поэтому действие медиатора на постсинаптическую мембрану прекращается. Образующийся холин активно поглощается пресинаптическими окончаниями и здесь, конденсируясь с ацетил-коферментом А, с помощью фермента холинацетилтрансферазы (ХАТ) образует новую молекулу ацетилхолина, поступающую в синаптическую везикулу. АХЭ инактивируется так называемыми нервно-паралитическими отравляющими веществами (зарин, зоман, w-газы), а также инсектицидами. В этом случае ацетилхолин накапливается в синапсах. При этом либо не происходит реполяризация постсинаптической мембраны, либо (во многих синапсах) наступает инактивация ХР, и постсинаптические каналы остаются постоянно закрытыми, хотя ацетилхолин присутствует в синаптической щели. В обоих случаях нарушается деятельность межнейрональных и нервно-мышечных синапсов и быстро наступает смерть - как правило, вследствие паралича дыхательных мышц.