

Распространение нервных импульсов. Сальтаторное проведение.
Проведение возбуждения по нервному или мышечному волокну осуществляется за счет локальных токов между возбужденными и покоящимися участками мембраны. При нанесении точечного раздражения на нервное волокно в этом участке возникает ПД. Внутренняя сторона мембраны в данной точке оказывается заряженной положительно по отношению к соседней, находящейся в невозбужденном состоянии. Между точками волокна, имеющими различный потенциал, возникает локальный ток, направленный от возбужденного (+ внутри мембраны) к невозбужденному (- внутри мембраны) участку волокна. Локальный ток оказывает деполяризующее влияние на мембрану в покоящемся участке и при достижении пороговой деполяризации в данном участке возникает ПД. Процесс генерации ПД последовательно распространяется по всему волокну.
Скорость проведения импульса будет тем выше, чем больше расстояние которое локальный ток, направленный, как уже говорилось, от возбужденного к невозбужденному участку волокна пройдет, сохраняя способность деполяризовать мембрану до порогового уровня. Это расстояние в свою очередь зависит от кабельных свойств аксона и будет тем больше, чем меньше электрическая емкость мембраны, больше ее поперечное сопротивление (меньше ток утечки) и меньше продольное сопротивление цитоплазмы. Сопротивление единицы длины аксона определяется площадью поперечного сечения аксона, и следовательно с увеличением диаметра (толщины) аксона скорость проведения импульсов возрастает. У кальмара и многих других беспозвоночных для быстрой передачи сигналов (до 100 м/с) в ходе эволюции появились аксоны огромного диаметра (гигантские аксоны - излюбленный объект исследования нейробиологов).
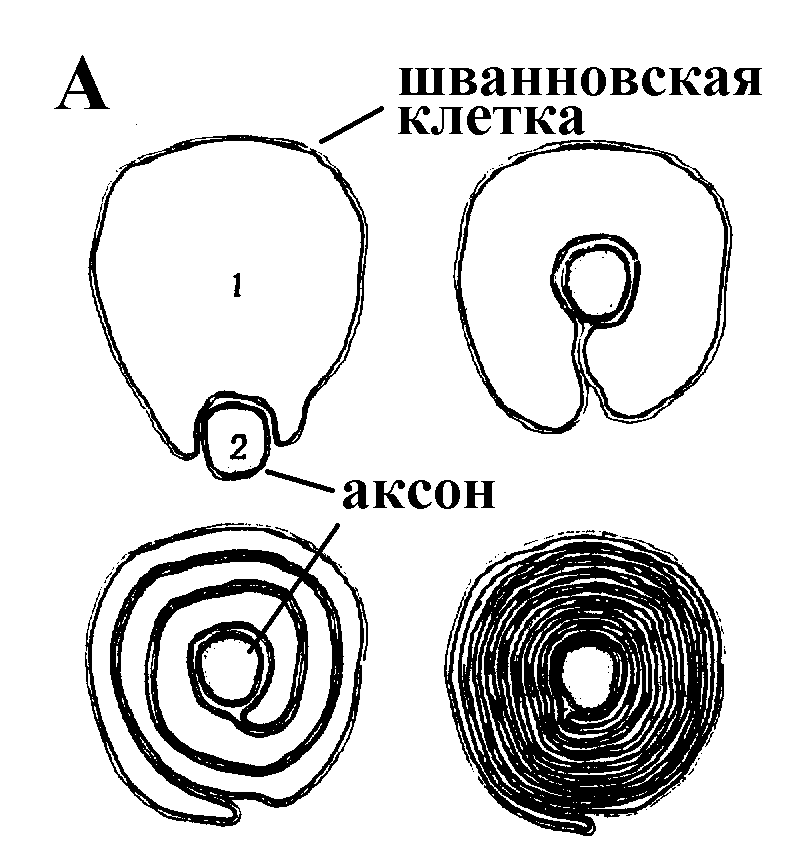
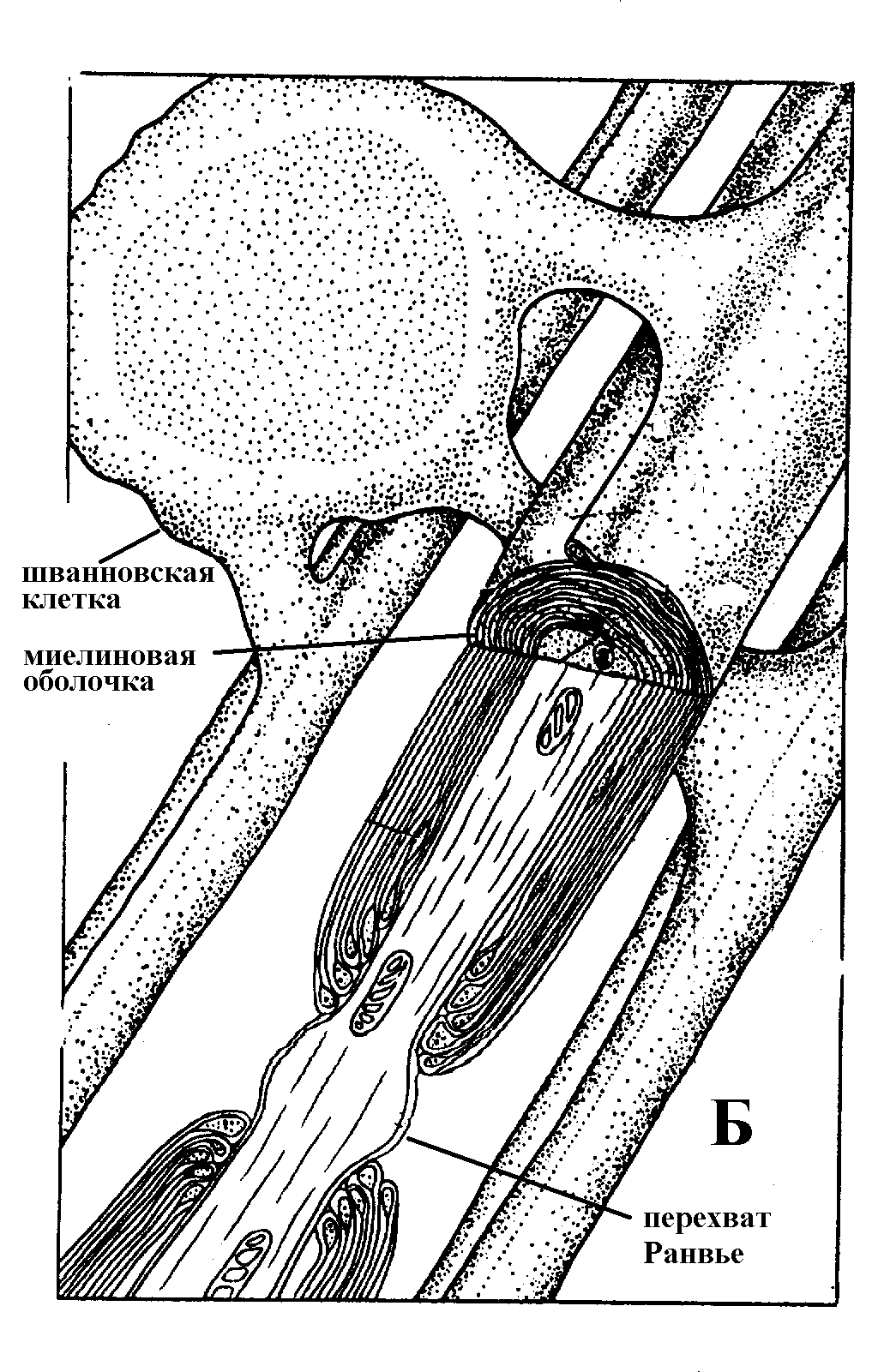
Рис. 13 А – образование миелиновой оболочки; Б – строение миелинизированного нервного волокна
Однако у позвоночных столь же высокая скорость проведения сигналов достигается гораздо более экономичным способом. Аксоны их нервных клеток заключены липопротеидную, миелиновую оболочку, которая по происхождению является плазматической мембраной особых клеток глии, называемых шванновскими клетками. В процессе формирования нервной системы, швановские клетки многократно накручиваются на аксон (до 300 спиральных слоев), при этом цитоплазма почти полностью выдавливается из межмембранного пространства (рис. 13). Изолирующий слой, образуемый миелиновой оболочкой, резко уменьшает емкость мембраны и утечку через нее тока.
Емкость мембраны
Мембрана клетки не только проводит ионные токи, но и накапливает заряд на своей внешней или внутренней поверхности. С точки зрения теории электричества, разделение зарядов на мембране означает, что мембрана обладает свойствами конденсатора. В обшем виде конденсатор состоит из двух проводящих пластин, отделенных друг от друга изолирующим материалом; в промышленных конденсаторах проводящие пластины обычно сделаны из металлической фольги, а изолирующая прослойка между ними — из пластика. В случае нервной клетки проводниками являются два слоя жидкости, находящиеся по обе стороны мембраны, а сама мембрана играет роль изолирующей прослойки. При зарядке конденсатора от батареи на одной из пластинок накапливается положительный заряд, в то время как на второй пластинке создается равный по величине запас отрицательного заряда. Емкость конденсатора (С) определяется количеством заряда (Q), запасаемым на один вольт потенциала (V), приложенного к пластинам конденсатора: С = Q/V. С измеряется в кулонах, деленных на вольт, т. е. в фарадах (Ф). Чем ближе друг к другу расположены пластины, тем более эффективно конденсатор способен разделять и накапливать заряд. Поскольку толщина мембраны клетки всего 5 нм, она способна накапливать достаточно большой заряд. Обычно емкость мембраны нервных клеток составляет 1 мкФ/см2. Преобразовав выражение, получаем Q = CV. При потенциале покоя —80 мВ, количество избыточного отрицательного заряда на внутренней стороне мембраны составит (1 · 10–6) х (80 ·10–3) = 8· 10–8 кулонов, деленных на см2, что соответствует 5 · 1011 одновалентных ионов (0,8 пмоль) на квадратный сантиметр мембраны.
Величину тока, протекающего внутрь конденсатора или из него, можно подсчитать на основе соотношения заряда и напряжения, учитывая, что ток (i, в амперах) есть скорость изменения заряда во времени, т.е. 1 ампер = (1 кулон)/(1 с). Поскольку Q = C/V, получим:
![]()
Скорость изменения заряда на конденсаторе прямо пропорциональна величине тока. Если ток постоянен, то потенциал будет меняться с постоянной скоростью dV/dt = i/C.
Соотношение тока и напряжения в цепи, содержащей резисторы (сопротивления) и конденсаторы (емкости), соединенные параллельно, проиллюстрировано на рис. 7.2. Прямоугольный скачок тока величиной i, приложенный к резистору (R), создает скачок напряжения на резисторе величиной V = iR (рис. 7.2А). Если тот же скачок тока приложить к конденсатору (С), то напряжение на конденсаторе будет накапливаться со скоростью dV/dt = i/C (рис. 7.2В). Когда эти два элемента, резистор и конденсатор, соединены параллельно (рис. 7.2С), то весь ток пойдет сначала на зарядку конденсатора со скоростью i/С; однако, как только на конденсаторе накопится какой-то заряд, ток потечет и через резистор. По мере нарастания тока, все большая его часть будет проходить через сопротивление, потому что скорость зарядки конденсатора будет постепенно снижаться. В конце концов весь ток будет течь через резистор, создавая на нем потенциал V = iR, a конденсатор будет полностью заряжен. По завершении скачка тока заряд из конденсатора постепенно рассеется на резисторе, а напряжение вернется к нулю. В нервной системе позвоночных нервные волокна большего диаметра обычно миелинизированы. На периферии миелин образуют шванновские клетки, а в ЦНС — олигодендроциты. Эти клетки плотно облегают нейроны, обвиваясь вокруг них. Мембраны так плотно соприкасаются друг с другом, что цитоплазма выдавливается из зазора между ними, в результате чего образуются тесно упакованные спиральные мембранные «обертки». Количество оберток (ламелей) варьирует от 10-20 до 160). Наличие 160 ламелей означает, что между плазматической мембраной аксона и внеклеточной жидкостью последовательно располагается 320 мембран. Таким образом, эффективное сопротивление мембраны возрастает в 320 раз, и во столько же раз снижается мембранная емкость. Миелин составляет обычно от 20 до 40 % общего диаметра волокна. Миелиновая оболочка прерывается через равные промежутки так называемыми перехватами Ранвье, в которых собственная мембрана аксона не покрыта миелином. Расстояние между перехватами обычно в 100 раз превосходит внешний диаметр волокна и составляет от 0,2 до 2 мм.
Миелиновые оболочки нервного волокна не сплошные, а прерываются вдоль волокна промежутками, которые называют перехватами Ранвье. Эти участки шириной всего около 0,5 мкм являются центрами электрической активности. Почти все натриевые каналы аксона сосредоточены в перехватах, где плотность каналов достигает несколько тысяч на 1 мкм2, тогда как в участках, покрытых миелиновой оболочкой, их почти нет. Поэтому изолированные участки мембраны не способны возбуждаться, но обладают превосходными кабельными свойствами - низкой емкостью и высоким сопротивлением для утечки тока. Благодаря этому локальные токи, обусловленные ПД в области перехвата Ранвье, легко доходят (путем пассивного проведения - электротонически) до следующего перехвата, быстро деполяризуют мембрану и возбуждают очередной потенциал действия. Такое проведение, когда сигнал распространяется вдоль аксона, "перескакивая" с одного перехвата на другой, называется сальтаторным. Миелинизация дает два главных преимущества: обеспечивает быстрое распространение ПД и сберегает метаболическую энергию, так как активное возбуждение происходит лишь на небольших участках в перехватах Ранвье.