

Внемикросомальный метаболизм ксенобиотиков
Вне микросом метаболизм происходит в гиалоплазме, лизосомах, пероксисомах, митохондриях.
Важнейшей реакцией детоксикации является реакция конъюгирования субстрата.
Эта реакция необходима для образования нетоксичных и легко выводимых продуктов метаболизма. В результате происходит инактивация и повышение растворимости, что облегчает выведение образующихся соединений в составе мочи, жёлчи, кала.
По механизму реакции конъюгации делятся на 2 типа.
1 тип. Сначала активируются конъюгирующие вещества, т.е. биомолекулы, а затем они переносятся на ксенобиотики с образованием конъюгата. Этот тип реакций наблюдается во всех тканях организма. В 1 типе реакций конъюгации выделяются:
а) глюкуронидная конъюгация.
Источником остатков глюкуроновой кислоты является УДФ-глюкуроновая кислота. Глюкуронидной конъюгации подвергаются природные соединения (билирубин, стероидные гормоны, витамин D) и ксенобиотики. Ксенобиотики могут вступать в глюкуронидную конъюгацию, если они имеют или приобрели в ходе модификации гидроксильную, карбоксильную, аминогруппу (обычно у ароматического кольца) и SH-группу.
Реакция конъюгации протекает с участием УДФ-глюкуронилтрансферазы.
Схема реакции:
RXH + УДФ-C6H9O6 → RX- C6H9O6 + УДФ-H
Ксено- УДФ-глюкуроновая глюкуронид
биотик к-та ксенобиотика
Среди ксенобиотиков (лекарств и ядов) глюкуронидной коньюгации подвергаются фенолы, полифенолы, фенольные стероиды, ароматические амины.
б) сульфатная конъюгация осуществляется при помощи активной формы конъюгирующего вещества: 3'-фосфоаденозин-5'-фосфосульфат (сокращённо ФАФС). Он конъюгируется с эндогенными токсическими продуктами гниения белков в кишечнике: индолом, скатолом, фенолом, а также стероидными иодтиронинами, токоферролами и нафтохинонами. Чтобы подвергнуться сульфатной конъюгации ксенобиотики должны, как правило, иметь циклические структуры, содержащие свободные OH- и NH2- группы.
Реакция сульфатной конъюгации происходит с участием: специального фермента сульфотрансферазы.
Схема реакции:
RXH + ФАФ~SO3H → RX-SO3H + ФАФH
Ксено- (ФАФС) сульфат 3'-фосфоаденозин-5'-фосфат
биотик ксенобиотика
 NH2
NH2


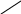
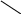 N
N

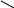

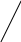 N
N
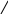


 O
O
N
N
O
O
N
N
ІІ ІІ
HO-S-O-P-O-H2C
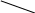
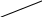


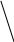
 ІІ
ІІ 5'
O
ІІ
ІІ 5'
O
O O
H H
3' H
H
H2O3PO OH
3'-фосфоаденозин-5'-фосфосульфат
в) ацетильная конъюгация
Источником подвижных ацетильных групп служит ацетил-SКоА, образовавшийся в ходе распада углеводов, ТАГ и аминокислот. Ацетилированию подвергаются природные вещества и ксенобиотики, имеющие свободную NH2-группу. Так обезвреживаются биогенные амины (серотонин и гистамин), ксенобиотики, имеющие NH2-группу у ароматического кольца, сульфаниламиды, гидразиды изоникотиновой кислоты, производные анилина. Фермент, катализирующий реакцию ацетилирования, ацилтрансфераза (ариламин-N-ацетилтрансфераза).
Схема процесса:
R-NH2 + CH3-CO~SCoA → R-NH-CO-CH3 + CoASH
г) метильная конъюгация – конъюгация с метильной группой, источником которой служит активная форма метионина: S-аденозил-метионин. Метилированию подвергаются ксенобиотики, содержащие NH2-группу или азот в гетероцикле, ОН-группу и SH-группу, т.е. происходит их N-, O- и S-метилирование. Так обезвреживаются моно- и полифенолы, пиридин, хинолин, изохинолин, тиоурацил. Фермент, катализирующий реакцию метилирования, - метилтрансфераза.
Схема процесса:
RXH + S-аденозилметионин → RX-CH3 + S-аденозилгомоцистеин
г) тиосульфатная конъюгация используется при ферментативном обезвреживании цианидов. Конъгирующим веществом является сера тиосульфата (или другие серосодержащие соединения).
Источником тиосульфата в тканях человека являются серусодержащие аминокислоты. Тиосульфатная конъюгация возможна с цианидами неорганического (синильная кислота, цианиды натрия и калия) и органического (ацетонитрил, акрилонитрил, бензилцианид, нитрил миндальной кислоты, малононитрил, циангидринглюкозиды и галогенцианиды) происхождения, если они освобождают в ходе гидролиза в организме цианид-ион.
Образующийся тиоционат менее токсичен, чем цианиды. Фермент, катализирующий реакцию тиосульфатной конъюгации, - тиосульфат-сульфидтрансфераза (роданеза - старое название).
Схема реакции:
CN- + S2O32- → SCN- + SO32-
Цианид тиосульфат тиоционат сульфит
2 тип. Сначала активируется ксенобиотик, который затем переносится на конъюгирующую биомолекулу с образованием конъюгата. Этот тип реакции наблюдается только в печени и в почках. Во 2 типе реакций выделяются:
а) глициновая конъюгация: субстратом конъюгации служат любые карбоновые кислоты, чаще циклические. Вначале происходит активирование субстрата, но не конъюгирующего вещества.
Схема процесса.


 СООН
СОSКоА
СООН
СОSКоА
+ АТФ + КоАSН → + АМФ + Н4Р2О7
пирофосфорная
кислота
Бензойная к-та бензоил-КоА
Фермент: ацетил-SКоА-синтетаза (арилацил-КоА-синтетаза).
Затем происходит конъюгация с NH2-группой глицина.
Схема процесса:
СО-SКоА + H2N-CH2-COOH → CO-NH-CH2-COOH + КоАSH
бензоилКоА гиппуровая к-та
Фермент: ацил-N-глицинтрансфераза, который специфичен только для ацилирования глицина, а не других аминокислот.
Аналогично происходит образование глициновых конъюгатов других соединений: ароматических кислот (никотиновой), стероидных кислот (холевой и дезоксихолевой) и др.
б) глутаминовая конъюгация.
Чаще наблюдается в ощутимых количествах у больных фенилкетонурией. У этих больных в больших количествах образуется фенилуксусная кислота, которая в печени и почках активируется с образованием фенилацетил-КоА и затем переносится на NH2-группу глутамина. При этом образуется конъюгат – фенилацетил-глутамин, выделяющийся с мочой.
Глутаминовая конъюгация ксенобиотиков (лекарств и ядов) в норме у человека не наблюдается.
Основными признаками повреждения при формирующихся патологических процессах в печени, имеющих диагностическое значение, являются:
повышение проницаемости мембран гепатоцитов;
снижение в них синтетических процессов;
увеличение размеров гепатоцитов, нарушение экскреции жёлчи;
нарушение процессов депонирования различных веществ;
ухудшение процессов дезактивации токсических метаболитов;
замещение клеток печени соединительной тканью;
нарушение синтеза различных белков.
В результате у больных можно наблюдать проявление следующих синдромов:
I. Синдром цитолиза (синдром нарушения целостности гепатоцитов) обусловлен нарушением проницаемости и деструкцией мембран гепатоцитов и их органелл, что приводит к выделению составных частей клеток в межклеточное пространство и кровь, в том числе и ферментов. Гиперферментемия развивается при острых поражениях печени (вирусных, токсических), при хронических аутоиммунных гепатитах, циррозах печени.
Для синдрома цитолиза характерно: повышение активности АЛТ, АСТ, альдолазы, глутаматДГ и других ферментов, а также гипербилирубинемия с повышением прямой фракции билирубина.
II. Синдром холестаза определяется нарушением оттока жёлчи. Если нарушается выделение жёлчи гепатоцитами (в случае поражения паренхимы печени: первичный билиарный цирроз и гепатиты различной этиологии), то развивается внутрипеченочный холестаз.
Закупорка жёлчных протоков, является причиной внепечёночного холестаза. Наиболее выражен холестаз при подпеченочной желтухе, которая развивается вследствие частичной или полной непроходимости общего жёлчного протока. Это приводит к застою жёлчи, повышению давления внутри протоков, повышению проницаемости, к диффузии жёлчи в печёночные клетки, лимфатическое пространство и кровь. Для синдрома холестаза типично повышение активности щелочной фосфатазы. Отмечается гиперхолестеринемия, ↑ уровня фосфолипидов, жёлчных кислот, ↑ прямого билирубина.
III. Синдром печёночно-клеточной недостаточности характеризуется снижением синтетических процессов, нарушением процессов депонирования различных соединений, дезактивации токсических метаболитов. Снижается уровень общего белка крови, особенно альбуминов. Такой синдром может быть при острых и обострении хронических гепатитов. Однако, при обострении хронических гепатитов ↑ фракция γ-глобулинов. При синдроме печёночной недостаточности понижается уровень I, V и VII факторов свёртывания крови, изменяется протромбиновое время. Повышается содержание NH3 в крови, фенолов, аминокислот; понижается уровень холинэстеразы.
IV. Воспалительный синдром обусловлен активированием клеток ретикулоэндотелиальной системы (РЭС). Для него характерно увеличение содержания в крови белков «острой фазы», а также диспротеинемия. Так, повышение α1, α2, β-глобулиновых фракций – это показатель воспалительного процесса при острых заболеваниях печени. Повышение γ-глобулиновой фракции обусловлено усиленным синтезом антител. При развитии иммунного ответа при остром воспалении наблюдается увеличение компонентов системы комплемента. Повышается синтез С-реактивного белка, необходимого для опсонизации возбудителей и активации комплемента.