

IV. Обезвреживающая функция печени
Печень – главное место обезвреживания токсических веществ как экзогенного, так и эндогенного происхождения. В печени осуществляется инактивация гормонов, лекарств, продуктов азотистого обмена, продуктов гниения в толстом кишечнике, обезвреживание билирубина.
Превращение ксенобиотиков происходит в разных органоидах клеток печени. В зависимости от локализации метаболизм ксенобиотиков делится на микросомальный и внемикросомальный.
Реакции внемикросомального метаболизма делятся на 2 типа.
К I типу относятся:
1) глюкуронидазная конъюгация;
2) сульфатная конъюгация;
3) ацетильная конъюгация;
4) метильная конъюгация;
5) тиосульфатная конъюгация.
Ко II типу относятся:
1) глициновая конъюгация;
2) глутаминовая конъюгация.
Микросомальный метаболизм ксенобиотиков.
Наиболее мощная система метаболизма находится в эндоплазматической сети (в микросомах). Микросомы являются фрагментами эндоплазматической сети. Микросомальное окисление служит универсальной биологической системой окисления неполярных соединений любого происхождения, поскольку цитохром Р450, непосредственно участвующий в окислении, находится в липидном бислое мембран. Субстрат, окисляемый цитохромом Р450, должен отвечать одному требованию – быть неполярным, т.е. проявлять специфичность не к структуре субстрата, а к его физико-химическим свойствам. Функциональная роль митохондриального и микросомального окисления различна.
Митохондриальное окисление – механизм использования кислорода в биоэнергетических процессах (синтез АТФ). В митохондриальном окислении играют роль реакции дегидрирования, а кислород является конечным акцептором электронов и используется для образования воды.
В процессах микросомального окисления активированный кислород непосредственно внедряется в окисляемое вещество. В микросомах находятся ферментные цепи окисления веществ. Они представлены двумя короткими цепями переноса электронов и протонов, встроенными в мембраны эндоплазматической сети или микросом.
Одна из них – монооксигеназная цепь окисления, в которой источником электронов и протонов является восстановленный НАДФН2. Источником НАДФН2 для монооксигеназной цепи является пентозофосфатный путь.
Другая – редуктазная цепь окисления, в которой источником ẽ и Н+ служит восстановленный НАДН+Н+. Источником НАДН+Н+ для редуктазной цепи является гликолиз.
НАДФН2 зависимая монооксигеназная цепь микросом состоит из:
фермент: НАДФ - Р450 -редуктаза;
флавопротеида (ФП2), коферментами которого служат ФАДН2 и ФМНН2;
цитохрома Р450.
Флавопротеид обдадает НАДФН2 дегидрогеназной активностью, причём ФАД+ и ФМН+ акцептируют 2Н+ и 2ẽ. С флавопротеина (ФМНН2) электроны (ẽ) транспортируются на цитохром Р450, а протоны (Н+) в окружающую среду.
Цитохром Р450 – последнее самоокисляющееся звено в этой цепи. Он относится к гемопротеидам. Содержит простетическую группу гем и имеет участки связывания для кислорода и субстрата (ксенобиотика). Белковая его часть представлена одной полипептидной цепью. Цитохром Р450 способен образовывать комплексы с оксидом углерода СО. Такой комплекс имеет максимум поглощения при λ=450 нм. Отсюда и название данного цитохрома.
Цитохром Р450 выполняет двойную функцию:
он активирует кислород посредством переноса на него электронов (образуется О22-);
использует активированный кислород для:
а) окисления вещества (ксенобиотика и других соединений);
б) образования воды.
В результате один атом кислорода включается в окисляемое вещество (RO), а другой, связывая два иона Н+ из среды, входит в состав воды.
НАДН+Н+ - зависимая редуктазная цепь окисления содержится не только в мембранах микросом, она имеется в наружной мембране митохондрий, ядерной мембране и клеточной мембране эритроцитов. Редуктазная цепь относится к самым быстрым реакциям биологического окисления. Она содержит:
фермент НАДН-цитохром В5-редуктазу. Это двухдоменный белок. Его коферментом является ФАД;
флавопротеид (ФП2), коферментами которого служат ФАДН2;
цитохром В5 – гемсодержащий белок;
фермент: стеарил-КоА-десатуразу, которая катализирует образование двойных связей в жирных кислотах, перенося электроны на кислород с образованием воды.
Редуктазная цепь может обмениваться с монооксигеназной цепью электронами. Например, электроны с ФП2 и цитохрома В5 могут переходить на цитохром Р450 и использоваться в окислении субстратов.
При острых поражениях печени активируется работа микросомальной системы. При хронических заболеваниях угнетается цитохром Р450 – зависимое гидроксилирование.
Активность микросомальной системы увеличивается с возрастом. Так, человеческий плод и новорождённый более чувствительны к лекарственным препаратам, т.к. у них снижена детоксикационная функция печени.
Индивидуальная чувствительность к лекарствам связана с индивидуальной активностью монооксигеназ.
ПФ-путь


 ↓
НАДФН2
ФАД+
ФМН
Fe3+
Н2О
↓
НАДФН2
ФАД+
ФМН
Fe3+
Н2О
 ROH
ROH
ЦитР450 НАДФ-Р450
редуктаза
R
 НАДФ+
ФАДН2
ФМНН2
Fe2+
О2
НАДФ+
ФАДН2
ФМНН2
Fe2+
О2
↓
2Н+
г ликолиз
ликолиз
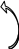 ↓
Н2О
↓
Н2О
Н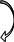
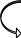


 АДН+Н+
ФАД+
Fe3+
АДН+Н+
ФАД+
Fe3+
R-OH

 цит
В5
НАДН
цитохром В5-редуктаза
цит
В5
НАДН
цитохром В5-редуктаза
стеароил КоА-десатураза
НАД+ ФАДН2 Fe2+
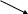
 О2
R
О2
R
2Н+
Рис.5. Схема монооксигеназной и редуктазной цепей окисления соединений в микросомах
Регуляция активности микросомальной системы осуществляется на уровне транскрипции или посттрансляционных изменений. Индукция синтеза позволяет увеличить количество фермента в ответ на поступление или образование в организме веществ, выведение которых невозможно без участия микросомального окисления. К числу индукторов относятся барбитураты, спирты, кетоны, стероидные гормоны, полициклические ароматические углеводороды. Например, фенобарбитал является индуктором в системе конъгации билирубина (↑ синтез ферментов микросомального и внемикросомального окисления). Следовательно, его введение ↓ концентрацию непрямого билирубина в крови. Поэтому небольшие дозы фенобарбитала дают новорожденным, чтобы уменьшить проявление патологической желтухи новорожденных.
У людей, страдающих хроническим алкоголизмом, активность ферментов микросомальной системы повышена, в связи с этим, при приёме барбитуратов нет ожидаемого эффекта, т.к. они быстро окисляются и выводятся из организма.
Существуют соединения, которые тормозят активность микросомального окисления. Например, два лекарственных препарата, принятых одновременно, могут конкурировать за ферменты микросомального окисления. В результате окисление одного из лекарств может снижаться, что приводит к накоплению неокисленного другого препарата. Единичный приём больших доз этанола тормозит микросомальное окисление.