

Морфологічною і функціональною одиницею цнс є нейрон.
А) Нейрон має:
Тіло – має оболонку(мембрану), ядро з ядерцем, цитоплазму з усіма органелами.
Дендрити – відносно короткі відростки, які сприймають інформацію. Поверхня їх вкрита великою кількістю шипиків (40000), які є місцем синаптичних контактів.
Аксон з пресинаптичними закінченнями. Аксон – довгий відросток, за допомогою якого передається інформація нервової клітини до інших відділів мозку або до робочого органа. Аксон має оболонку, початковий сегмент, аксоплазму, колатеральні гілки та телодендрон. Початковий сегмент знаходиться між аксонним горбком і початком оболонки нервового волокна. На ньому можуть закінчуватися аксони інших нейронів, тоді збудження передається по цьому аксону, минаючи клітину. Колатералі – це відведення аксона, по яких розповсюджується одні й ті самі нервові імпульси на інші нейрони. Телодендрони – місця утворення синапсів.
Аксонний горбок – це ділянка нейрона, де починається аксон. Це найбільш збудлива частина клітини, саме тут виникає потенціал дії нервових клітин, що розповсюджується до аксона.
Класифікація нейронів.
І. За кількістю та характером відростків нейрони поділяються на:
Уніполярні (одновідросчасті) – зустрічаються в вузлових нервових системах безхребетних. Псевдоуніполярні (розгалужений відросток).
Біполярні (двовідросчасті) – чутливі нейрони, нюхові нейрони, нейрони сітківки ока, слухові нейрони.
Мультиполярні (багатовідросчасті).
Аксони разом з оболонками, що входять до складу периферичних нервів, називають нервовими волокнами (мієлінові та безмієлінові). Всередині волокна міститься осьовий циліндр з нейрофібрилами. Нервові волокна здатні до регенерації. Збудження по нервовим волокнам проводиться ізольовано в обох напрямках від місця його виникнення. З функціональної точку зору нервові волокна характеризуються високою збудливістю, лабільністю і відносною невтомою.
За будовою, швидкістю проведення збудження і тривалістю потенціалу дії нервові волокна поділяють на три групи:
група А – товсті (діаметр 4-20 мкм) мієлінові волокна з великою швидкістю проведення збудження (до 120 м/с);
група В – великі немієлінові волокна вегетативної нервової системи (діаметр 1-3 мкм, швидкістю проведення збудження 3-14 м/с);
група С – тонкі немієлінові волокна (діаметр 0,5 мкм) з малою швидкістю проведення імпульсу.
ІІ. За функціями розрізняють:
Аферентні (чутливі) нейрони – несуть інформацію від рецепторів до ЦНС. Вони біполярні (їх тіла знаходиться поза ЦНС -в гангліях спинного мозку або черепних нервів)
Еферентні (рухові) нейрони – посилюють нервові імпульси до робочих органів. Їх тіла (окрім ВНС) знаходяться в ЦНС.
Проміжні нейрони – передають інформацію від одного нейрона до іншого, здійснюють попередній аналіз. Утворюють нейронні сітки. За функцією вони бувають: збудливі (активують інші нейрони) і гальмівні (знижують активність інших нейронів.
Головними функціями нейрону є: генерація процесу збудження, передача його на інші клітини або робочі органи, здійснення трофічних впливів на тканини, що іннервуються, синтез речовин-регуляторів, синтез метаболітів, необхідних для росту і регенерації самого нейрону і його відростків.
Б) Нейроглія – клітини різної форми, які супроводжують нервові клітини. Відростки клітин глії переплітаються між собою і утворюють густу сітку, яка заповнює простір між нервовими клітинами і капілярами. Вони складають 40% об’єму мозку. З віком кількість нейронів зменшується, а нейроглії – збільшується.
Види:
Астроцити (60%) – беруть участь в утворенні гематоенцефалічного бар’єру, обміну медіаторів ЦНС, в імунних реакціях мозку та ін..
Олігодендроцити (25-30%) –утворюють мієлінову оболонку нейронів. На периферії цю функцію виконують швановські клітини. Вони також беруть участь в імунних реакціях мозку (поглинають мікроорганізми).
Епендимні клітини – вистеляють шлуночки головного мозку. Беруть участь в утворенні гематоенцефалічного бар’єру і процесах секреції спинномозкової рідини.
Мікроглія (10%) – бере участь в фагоцитозі.
В) Нервові центри (анатомічні) – це сукупність нейронів, розташованих в ЦНС. НЦ забезпечують рефлекторну діяльність, мають велику кількість нейронів і синапсів.
Фізіологічне розуміння нервового центру – це складне функціональне об’єднання декілька анатомічних нервових центрів, розташованих на різних рівнях ЦНС і обумовлюють за рахунок своєї активності складні рефлекторні акти (н-д, за травлення відповідають багато нервових центрів на різних рівнях ЦНС – функціональне об’єднання нервових центрів).
Властивості нервових центрів:
Однобічність проведення збудження – від рецептора до ефектора, що пояснюється «клапанною» дією синапсів.
Затримка проведення збудження (через велику кількість синапсів) – пов’язано з процесами виділення медіатора, хімічними процесами, які відбуваються в синапсі, з виникненням ЗПСП і регенерацією ПД (1,5-2 мс). Час від подразнення до рефлекторної відповіді зветься латентним (прихованим) періодом рефлексу -ЛП. Він прямо пропорційно залежить від кількості нейронів даного рефлексу (н-д, колінний моносинаптичний рефлекс має ЛП 24 мс, а зорова реакція – 200 мс.
Сумація збудження – виникає або при нанесенні слабких, але ритмічних подразнень, або при одноразовій дії декількох підпорогових подразників. Механізм дії пов’язаний з накопиченням медіатора на постсинаптичній мембрані і підвищенням збудливості клітин нервового центру.
Трансформація ритму збудження – частота імпульсації еферентного нейрону ніколи не відповідає частоті аферентного сигналу. Це пов’язано з рядом особливостей як синаптичних передач між нейронами, так і самих клітин нервового центру.
Післядія збудження (рефлекторна післядія) – після припинення подразнення збудження в ЦНС може продовжуватись ще деякий час. Вона може бути:
А) короткочасною (1-3 мс) – пов’язана зі слідовою деполяризацією оболонки клітини;
Б) тривалою (секунди) – зумовлена циркуляцією збуджень по нервовим ланцюгам нервового центру (збудження циркулює і підтримує саме себе – реверберація). Циркуляція нервових імпульсів в НЦ буде продовжуватися до тих пір, поки не настане втомлення синапсів або коли прийдуть гальмівні імпульси.
6) Тонус НЦ – завдяки реверберації НЦ завжди знаходяться в стані тонусу.
Вони спонтанно посилюють імпульси до робочого органу без аферентних
впливів. Н-д, тонус НЦ забезпечує тонус м’язових волокон, стінок
кровоносних судин.
7) Полегшення збудження – при наявності імпульсів один за одним або коли
збудження нейрона виникає внаслідок збудження близько розташованих
синапсів. Кожний наступний імпульс потребує меншого критичного рівня
деполяризації.
Ритмічна активність НЦ – форма підвищення готовності до відповідних
реакцій. В НС є нейрони, які проявляють фонову (спонтанну) активність –
мимовільне виділення медіатора, наслідком чого є спонтання деполяризація,
реверберація імпульсів.
Стомлюваність НЦ - через деякий час зменшується сила рефлекторної
реакції, і навіть припиняється. Пов’язане з порушенням передачі в синапсах
і виснаженні запасів медіаторів та ін..
10) Чутливість НЦ до хімічних речовин і нестачі кисню.
А) Синапс – місце контакту двох нейронів. За допомогою синапсу відбувається передача нервового імпульсу від одного нейрону до інших, утворюються нейронні сітки. Кожен нейрон має багато синапсів – від 5000 до 200000 синапсів (на мембрані тіла немає вільної ділянки від синапсів).
Класифікація центральних синапсів.
І. За розташуванням:
Аксосоматичні – аксон одного нейрону розташований на тілі іншого.
Аксодендритичні – аксон на дендриті.
Аксоаксональні - - аксон на аксоні.
Дендродендритичні – дендрит на дендриті.
Дендросоматичні – дендрит на тілі.
ІІ. За функціями:
1) Збудливі – коли нервовий імпульс, прийшовши до цього синапсу, збуджує
постсинаптичний нейрон. Під впливом нервових імпульсів вивільняється
збуджувальний медіатор (ацетилхолін, норадреналін), який через синаптичну
щілину потрапляє до постсинаптичної мембрани і викликає короткочасне
підвищення її проникності для іонів натрію і виникнення деполяризації.
Коли деполяризація досягає критичного рівня, виникає розповсюджене
збудження – потенціал дії.
2) Гальмівні – коли імпульс приводить до пригнічення активності
постсинаптичного нейрону. В цих синапсах виділяються гальмівні медіатори
(ГАМК).
Вони змінюють проникність постсинаптичної мембрани по
відношенню до іонів калію або хлору. В результаті підвищується рівень
мембранного потенціалу (гіперполяризація), що затримує подальше
розповсюдження збудження.
ІІІ. За механізмом передачі сигналу.
Хімічні – більшість синапсів ЦНС. За будовою нагадують периферичний
нервово-м’язовий синапс.
2) Електричний – в цих синапсах синаптична щілина вужа за химичний
синапс. Має прямі контакти окремих ділянок пре- і постсинаптичної ділянок.
Нервовий імпульс передається швидко, без синаптичної затримки. Синапси
мають двобічну провідність, малу втомлюваність.
Б) Медіатори ЦНС. До них відносять:
Аміни (ацетилхолін, норадреналін, адреналін, дофамін, серотонін).
Амінокислоти (гліцин, глютамін, аспарагінова кислота, ГАМК)
Пурінові нуклеотиди (АТФ).
Нейропептиди (гіпоталамічні ліберини і статини, опіоїдні пептиди, вазопресин, гастрит та ін.).
В багатьох нейронах може бути більш одного медіатора.
За механізмом дії медіатори бувають:
Іононотропні – при взаємодії з рецепторами постсинаптичної мембрани змінюють проникність іонних каналів.
Метаботропні – вплив на постсинаптичну мембрану оказують через активацію специфічних ферментів постсинаптичної мембрани. В мембрані активуються вторинні посередники – месенджери, які запускають каскади ферментативних реакцій в клітині або мембрані. Ці метаболічні зміни достатньо глибокі і можуть викликати зміни генома клітини, беруть участь в формуванні пам’яті.
3. Механізм передачі збудження в центральному хімічному аксосоматичному синапсі.
ПД поширюється по мембрані аксона.
ПД розповсюджується по пресинаптичній мембрані.
Підвищується проникність пресинаптичної мембрани для іонів кальцію.
Вхід іонів кальцію в нервове пресинаптичне закінчення за градієнтом концентрації (пасивно).
Вихід медіатора в синаптичну щілину.
Дифузія медіатора до постсинаптичної мембрани.
Взаємодія з мембранними циторецепторами.
Збільшення проникності постсинаптичної мембрани для іонів натрію.
Вхід іонів натрію в тіло клітини через постсинаптичну мембрану
Деполяризація мембрани – збуджуючий постсинаптичний потенціал (ЗПСП місцевого збудження – здатний до сумації і не здатний до розповсюдження). ЗПСП має підпороговий рівень, і тому недостатній для ПД на постсинаптичній мембрані. Генерація ПД виникає в результаті сумації ЗПСП. Цьому сприяє здатність ЗПСП поширюватися на сусідні ділянки мембрани.
ЗПСП поширюється на сусідні ділянки постсинаптичної мембрани та мембрани
аксонного горбику з допомогою місцевих струмів. Ці струми в незбуджених
ділянках мембрани викликають деполяризацію мембрани.
Особливості передачі збудження через хімічні синапси:
Однобічне проведення.
Сповільнення проведення – характеризується наявністю синаптичної затримки
Швидке порушення проведення через виснаження запасів медіатора.
Один ПД не проходить через центральний синапс. Це виникає тому, що на постсинаптичній мембрані один ЗПСП має амплітуду 1мВ та тривалість 15 мс. Оскільки поріг деполяризації аксонного горбика складає 10-15 мВ, один ЗПСП, що поширився на мембрану аксонного горбика, не може викликати деполяризацію цієї мембрани до критичного рівня.
Умовою передачі збудження через центральний нервовий синапс є сумація ЗПСП на тілі нейрона: просторова сумація (на тілі нейрона одночасно функціонує декілька збудливих синапсів) і часова сумація =послідовна (кожен наступний ЗПСП починається тоді, коли ще не скінчився попередній; частота ПД має бути не менше 66 Гц).
Для збудження нейрона виникнення ЗПСП в одному синапсі недостатньо. Необхідно, щоб збудження виникло одночасно не менш ніж у 50 пресинаптичних утвореннях. Лише в цьому випадку ЗПСП досягає критичного рівня. Найбільша збудливість у перетинці початкового сегмента аксона. Саме тут виникає ПД (вже при деполяризації 10 мВ), який потім розповсюджується по аксону на тіло клітини.
Останні дві особливості відносяться тільки до центральних синапсів.
4. Гальмівні синапси (механізми центрального гальмування). Пре- і постсинаптичне гальмування.
Гальмування – це активний нервовий процес, який проявляється у пригніченні збудження або в повному його припиненні, у зменшенні фізіологічної реакції організму або припиненні її.
Форми гальмування:
1) Первинне - необхідна наявність спеціальних гальмівних структур (гальмівних нейронів і гальмівних синапсів). Гальмування виникає первинно, без попереднього збудження: пресинаптичне і постсинаптичне.
А)Пресинаптичне гальмування.
Морфологічна основа – аксо-аксональні синапси: аксон проміжного гальмівного нейрона утворює синапс на аксоні збудливого нейрона . Виникає зменшення або блокада виділення медіатора в синаптичну щілину, який передає збудження. В основі – зміни пресинаптичної мембрани ( повільна і тривала деполяризація пресинаптичної мембрани).
Б) Постсинаптичне гальмування ( зниження збудливості оболонки нейрона, його дендритів) – основний вид гальмування в ЦНС. Пов’язаний з діяльністю спеціальних гальмівних нейронів.
Гальмівний синапс знаходиться між збудливим синапсом і аксонним горбком. Основний медіатор – ГАМК, гліцин.
Стимул викликає на гальмівному синапсі гіперполяризацію постсинаптичної мембрани ( а не деполяризацію) –виникає гальмівний постсинаптичний потенціал: ГПСП.
Механізм постсинаптичне гальмування (на прикладі аксосоматичного синапса):
В основі гіперполяризації - підвищена проникність мембрани для калія і для хлора. Відкриття каналів для хлору призводить до пасивного транспорту всередину гальмівного медіатора іонів хлору. Ці додаткові негативні іони і збільшують заряд мембрани. Гальмівной медіатор відкриває канали для калію – калій виходить з нейрону (також сприяє гіперполяризації).
Боротьба між ГПСП і ЗПСП - ГПСП перешкоджає досягненню в гальмівному медіаторі критичного рівня деполяризації (ЗПСП) під впливом збуджувальних імпульсів –знижуеться збудливість клітин.
2)Вторинне – розвивається в результаті зміни функціональної активності звичайних збудливих нейронів.
Лекція №1.
Тема: Фізико – хімічні властивості крові.
План: 1. Поняття про систему крові. Склад і об’єм крові. Основні функції крові.
Гематокритний показник.
2.Основні фізіологічні константи крові.
3. Плазма, склад, роль білків плазми.
4. Осмотичний і онкотичний тиски. Регуляція сталості осмотичного тиску.
5. Кислотно – основний стан крові. Буферні системи. Види. Механізми функціонування.
І. Кров – це різновид сполучної тканини. Вона заповнює кровоносні судини. Складається з міжклітинної речовини – плазми (55-60%) й клітин крові або формених елементів (40-45%). Гематокрит – це співвідношення формених елементів і плазми крові ( у чоловіків 0,41 – 0,53, у жінок 0,36-0,46); він зменшується при анеміях, а збільшується при збільшенні клітин крові (пухлини крові і нирок, дегідратації).
Складові частини крові постійно руйнуються і замінюються новими. Функціонує спеціальна система, де відбувається руйнування і утворення складових частин крові. Кров і кровотворні органи (кістковий мозок, селезінка, печінка, лімфатичні вузли) утворюють цілісну фізіологічну систему – систему крові. В організмі людини кров становить 6 -7, 7 % маси тіла, тобто для дорослої людини це -4,5 – 6 літрів. Об’єм циркулюючої крові у судинному руслі (ОЦК) є однією з констант організму. Проте ОЦК не постійна величина. Він залежить від віку, статі, функціонального стану органів конкретної людини, у новонароджених ОЦК становить 10%, а у період статевого дозрівання поступово досягає рівня, як у дорослих. У людей, які ведуть малорухомий спосіб життя, ОЦК нижчий, а у фізично тренованих, навпаки, вищий від середнього рівня. Так, у спортсменів, здатних витримувати велике фізичне навантаження, ОЦК досягає 10% маси тіла.
У стані фізіологічного спокою по кровоносних судинах циркулює тільки половина всієї крові, а остання знаходиться у депо крові (печінці, селезінці, легенях, судинах шкіри). При нестачі кисню в організмі клітини крові виходять з депо і поповнюють кровоносні судини, підвищуючи цим киснево – транспортну функцію крові. Об’єм крові досить сталий. При крововтрата він швидко вирівнюється внаслідок виходу рідин з тканинного простору в кров, але при значних кровотечах, коли втрачається 1/3 крові, може настати смерть. У цих випадках необхідне швидке переливання крові або рідини, що її замінює.
Оскільки у кров потрапляють усі продукти життєдіяльності організму, а її морфологічний склад залежить від співвідношення процесів утворення і руйнування, то її хімічний і морфологічний склад є одним з найбільш чутливих показників стану організму при виникненні різних захворювань.
Основні функції крові:
Дихальна функція полягає у зв’язуванні і перенесенні 02 від легень до тканини і СО2 – із тканин до легень.
Трофічна функція крові пов’язана із забезпеченням усіх клітин організму поживними речовинами, що надходять із органів травлення або інших органів.
Екскреторна функція – кров транспортує з тканин кінцеві продукти метаболізму до органів виділення.
Терморегуляторна функція – кров несе тепло від енергоємних органів та зігріває органи, що втрачають його.
Забезпечення водно-електролітного обміну: в артеріальній частині більшості капілярів рідина і електроліти надходять у тканини, у венозній – вони повертаються у кров.
Захисна функція – кров забезпечує захист організму від бактерій, вірусів і генетично чужерідних клітин і речовин.
Гуморальна регуляція – кров переносить гормони та інші біологічно активні сполуки від клітин, де вони утворюються, до інших органів та тканин. Цим самим забезпечується хімічна взаємодія між усіма частинами організму.
Перераховані функції крові спрямовані на підтримання гомеостазу організму
ІІ.Основні фізіологічні константи крові.
До фізико-хімічних властивостей крові відносять:
1.Осмотичний тиск крові – 7,5 атм;
2. Реакцію крові – рН =7,36-7,4;
3. Щільність крові (густина) – вища від щільності води за рахунок розчинених у ній речовин. Густина плазми становить 1,025-1,034 г/см3, цільної крові – 1,05-1,06 г/см3, еритроцитів – 1,09 г/см3.
4. В’язкість крові – 5,0 (визначається відносно в’язкості води=1). В’язкість визначається тертям крайніх шарів плазми до стінки судин. Збільшує в’язкість крові вміст еритроцитів, білкових молекул. За рахунок в’язкості кров створює опір кровотоку.
ІІІ. Плазма. Плазма містить 90-92% води і 9-10% сухого залишку (органічні і неорганічні речовини). Вода постійно оновлюється ( за одну хвилину 70% води). Обмін води між плазмою і міжклітинним простором здійснюється у мікроциркуляторному руслі.
Органічні речовини крові:
- білки (альбуміни, глобуліни, фібриноген) – 7,2%;
- глюкоза (3,33 – 5,55 ммоль/л); це єдине джерело для мозку (при концентрації нижче 2,22 ммоль/л у людини виникає гіпоглікемічна кома – втрата свідомості, порушення кровообігу і дихання).
- амінокислоти;
- ліпіди;
- гормони;
- вітаміни;
- продукти проміжного обміну, сечовина, сечова кислота.
Білки плазми.
1. Альбуміни у руслі крові містяться тривалий час (10-15 діб). Їх функції: 1) транспортна функцію (тому що мають сумарно велику площу поверхні), 2) Резервні білки для утворення амінокислот; 3) створюють онкотичний тиск (обмін води між тканинами) – для регулювання об’єму крові( регулюють ОЦК та інтенсивність утворення сечі).
2. Глобуліни – група білків, яка поділяється на фракції: альфа, альфа, бета, гама-глобуліни. Циркулюють глобуліни 5 діб. Їх функції: 1) транспортна – альфа-глобуліни переносять ліпіди, тироксин, гормони наднирників, мідь; бета-глобуліни – залізо, гем, ліпіди,
2) захисна – гама-глобуліни є антитіла.
3. Фібриноген – найбільший білок плазми. Його функція – зсідання крові й припинення кровотечі.
Білки плазми мають буферні властивості (підтримують рН крови), забезпечують артеріальний тиск (внаслідок в’язкості), заважають осіданню еритроцитів, підтримують в’язкість крові (підтримують тиск в судинах).
Неорганічні речовини крові.
Мінеральні речовини становлять 0,9 % маси плазми. Тут переважно містяться натрій, калій, кальцій, хлор,НСО3 -,НРО4 2-. Деяких речовин знаходиться незначно – це мікроелементи (мідь, залізо). Більшість мінеральних речовин постійна, їх рівень регулюють гормони (кальцій).
ІУ. Осмотичний і онкотичний тиски. Регуляція сталості осмотичного тиску.
Осмотичний тиск – основан на осмосі (протягування деяких речовин до себе води) і дорівнює 7,5 атм. Осмотичний тиск (сила, з якою розчин спрямовується до речовини) визначається концентрацією розчинених молекул. 96% загального осмотиску плазми припадає на частку неорганічних електролітів (солей), серед яких основна роль належить NaCl (60-80%). У клінічній практиці використовують переливання кровозамінників. Для цього використовують ізотонічні (фізіологічні) розчини. Найпростіший кровозамінник – 0,9 % розчин натрію хлориду, який створює осмотиск, близький до 7,5 атм. Розчин, осмотиск якого перевищує осмотиск плазми, зветься гіпертонічним, а, якщо має менший осмотиск – гіпотонічним.
Онкотичний тиск – це частина осмотичного тиску, який створюється органічними речовинами плазми (білками). На білки плазми припадає 0,03-0,04 атм (25-30 мм рт.ст.) тиску. Завдяки вищому онкотичному тиску в крові утримується вода.
Осмотиск і онкотиск забезпечують водний обмін між середовищами організму, впливають на обмін води між плазмою крові і клітинами крові.
При порушенні осмотиску або онкотиску у плазмі змінюються функції клітин крові і тривалість їх життя. Так, при зниженні осмотиску вода буде надходити у клітини крові, виникає розрив оболонок клітин (осмотичний гемоліз). При підвищенні осмотиску вода виходить із клітин – зморщування клітин.
Регуляція осмотиску.
Активність біосинтезу білків в печінці;
Робота органів виділення – нирки, потові залози.
При ураженні нирок порушується виведення білків, виникають набряки тканин.
У. Кислотно – основний стан крові. Буферні системи. Види. Механізми функціонування.
Реакція крові (рН) зумовлена співвідношенням в ній водневих і гідроксильних іонів. Реакція крові є слабко лужною. В стані фізіологічного спокою рН арт. крові дорівнює 7,4, а у венозній – 7,36. рН крові жорстко запрограмовано – життя можливе лише при зміщенні у вузькому діапазоні рН: від 7,0 до 7,8. В процесі діяльності клітин у них постійно накопичуються кислі продукти обміну, які зменшують рН. Підтримання сталості рН крові на оптимальному рівні здійснюється певними системами – буферними.
Буферні системи:
1) Буферна система гемоглобіну (гемоглобін-калійна сіль Нв)– найпотужніша, становить 75% буферної місткості крові. У тканинах гемоглобін крові виконує функцію лугу і зв’язує СО2 і Н+, а в легенях – функціонує як кислота, запобігає закисленню крові після виділення з неї вуглекислоти.
2) Карбонатна БС( вугільна сіль Н2СО3, бікарбонат натрію NаНСО3)– утворюється слабкою вугільною кислотою і її сіллю – Na СО3. Якщо додати до такої речовини надлишок NаНСО3 – солі сильної кислоти, то вона витіснить слабку кислоту з її сполук і з лугу. В розчині залишається слабка кислота і сіль сильної кислоти. Якщо до буферної системи додати лугу, то утворюється сіль слабкої кислоти і вода. Тому в обох випадках зменшується відхилення активної реакції.
3) Фосфатна буферна система (одноосновний NаН2РО4 і двохосновний фосфат натрію Nа2НРО4) – утворюється дигідрофосфатом (NаН2РО4) і гідрофосфатом (NаНРО2). Перша сполука поводить себе як слабка кислота, друга – як луг.
4) Буферна система білків – зумовлена наявністю в їхньому складі кислотних і лужних груп. Тому в кислому середовищі білки дисоціюють як луги, зв’язуючи водневі іони, а в лужному зв’язують іони гідроксилу.
Збереженню сталості рН сприяє функціонування легень, нирок, шлунка, кишок, потових залоз. Через нирки та потові залози виводиться надлишок лугів і кислот.
Зміщення рН крові ближче до кислого середовища називають ацидоз, а до лужного – алкалоз.
Лекція № 3.
Тема: Захисна функція крові.
План: І. Лейкоцити. Кількість. Лейкоцитоз. Лейкопенія. Лейкоцитарна формула.
Функція різних видів лейкоцитів. Регуляція кількості лейкоцитів.
ІІ. Тромбоцити. Кількість. Функції.
ІІІ. Гемостаз. Види. Механізми розвитку. Коагулянти. Антикоагулянти.
І. Лейкоцити – повноцінні клітини, що містять ядро та органели, розміром 10-17 мкм. За формою ядра вони поділяються на 5 видів. В 1 л крові міститься 4-9 х 10 9 лейкоцитів. Збільшення їх кількості називається лейкоцитозом, а зменшення – лейкопеніею.
Лейкоцитоз поділяється на фізіологічний (м’язовий, травний) або патологічний (запалення, пухлини і т.п.). Місцем утворення лейкоцитів є червоний кістковий мозок, а в лімфатичних вузлах та селезінці вони дозрівають.
Лейкоцити виконують свої захисні функції переважно у сполучній тканині різних органів. У руслі крові лейкоцити циркулюють протягом кількох годин (4-72), потім вони виходять через стінку капілярів і розселюються по тканинах. У тканинах лейкоцити можуть перебувати протягом багатьох діб (8-10).
Властивості лейкоцитів:
Амебоподібна рухомість – лейкоцити завдяки псевдоніжки (виросток цитоплазми) можуть рухатися до патологічного фактора (хемотаксис).
Діапедез – проникнення крізь стінку капілярів.
Фагоцитоз – поглинання чужорідних речовин.
Лейкоцити поділяються на дві основні групи за властивостями цитоплазми:
Зернисті (гранулоцити) – мають в цитоплазмі дрібні зерна, які фарбуються різними фарбниками: нейтрофіли – нейтральними фарбниками у сине-фіолетовий колір, еозинофіли (ацидофіли) – кислими фарбниками у червоний колір, базофіли – лужними фарбниками у фіолетовий колір. Їх ядра теж різняться за формою: нейтрофіли мають паличкоподібні або сегментовані ядра (частіше 3-4 сегмента); еозинофіли – сегментовані ядра (2 сегмента); базофіли – сегментовані ядра без чіткого розділення на сегменти.
Незернисті (агранулоцити) – не мають в цитоплазмі зерен. Це моноцити (великі клітини з великим кулястим або бобоподібним ядром) і лімфоцити
(різних розмірів клітини з великим кулястим ядром).
Відсоткове співвідношення між окремими видами лейкоцитів зветься лейкоцитарною формулою. Визначення цієї формули має важливіше значення в клініці. Н-д, при запаленні збільшується кількість нейтрофілів, при глистах і алергіях – еозинофілів. При появі незрілих форм лейкоцитів – говорять про зсув формули ліворуч, при збільшенні зрілих форм лейкоцитів – праворуч.
Нейтрофіли – 65-75% (сегментоядерних – 45-72%, паличкоядерних – 1-5%).
Еозинофіли – 0,5-5 %
Базофіли – 0-1%
Моноцити – 3-11%
Лімфоцити – 18-37%.
Функції лейкоцитів:
Нейтрофіли:
а) захисна - фагоцитоз (мікрофаги). Травлення мікробів відбувається завдяки різних факторів в гранулах, лізосомних ферментів (фагоцитин, гідролази, поліпептидами). Нейтрофіли утворюють основу гною.
б) синтетична - утворення інтерферону (речовина, яка діє на віруси).
У руслі крові міститься незначна кількість зрілих клітин. У 20-40 разів більше їх є в органах-депо, зокрема кровотворних – кістковому мозку, селезінці, печінці, а також у капілярах легень. Після утворення зрілий нейтрофіл ще протягом 5-7 діб залишається в кістковому мозку. Звідси нейтрофіли можуть виходити і поповнювати пул циркулюючих клітин, що накопичилися біля місця ушкодження, в ділянці запалення.
Вміст нейтрофілов у крові може збільшуватися як за рахунок інтенсивного лейкопоезу, так і шляхом перерозподільної реакції, що виникає при емоціях, фізичному навантаженні, травленні, різноманітних стресах. Активний вихід лейкоцитів із кісткового мозку призводить до появи в руслі крові юних форм: паличко ядерних, і навіть мета мієлоцитів. Зростання їх кількості в крові свідчить про активізацію процесів творення і виходу нейтрофілів із кісткового мозку. Перерозподільний лейкоцитоз є наслідком не тільки виходу нейтрофілів з органів-депо, але й зміни засобу циркуляції. У звичайних умовах у тоці крові нейтрофіли діляться на дві частини – пул клітин, що циркулюють по центру судин, та пул пристінкових клітин. При вищезгаданих станах лейкоцити відходять від стінок і приєднуються до циркулюючих клітин.
2. Базофіли:
а) захисна - участь у запаленні (синтезують гістамін).
б) запобігають зсіданню крові –синтезують гепарин
в) синтетична - синтез БАВ і медіаторів: брадикінін, серотонін; вони беруть участь в алергічних реакціях. Тому базофілія є однією з ознак сенсибілізації організму при алергіях. Тобто базофіли сприяють розвитку запалення, а після ліквідації пат.процесу беруть участь у розсмоктуванні осередку запалення.
Еозинофіли:
а) захисна - фагоцитоз (мікрофаги) чужорідних білків і білкових токсинів.
б) адсорбують на своєї поверхні АГ і білкові токсини, потім їх руйнують.
в) попереджують розвиток алергій. Інактивують ряд субстанцій анафілаксії, зменшують виразність реакцій негайної гіперчутливості. Еозинофіли під впливом хемотаксичних факторів мігрують до місця появи невеликої кількості антигену, де відбувається реакція антиген-антитіло. Тут вони утворюють наче захисний вал у вигляді місцевого фіброзування, що затримує поширення даної реакції по організму.
г) цитотоксичний вплив на гельмінти та їх личинки (завдяки основного білка в гранулах).
Виходячи з кісткового мозку, еозинофіли у крові циркулюють недовго (2 г), після чого мігрують переважно у покривні тканини. Вміст їх у тканинах приблизно у 100 разів більший, ніж у кровотоці. При тривалому перебуванні у організмі гельмінтів, алергізації розвивається еозинофілія – збільшення кількості циркулюючих еозинофілів. Вона зумовлена виходом із кісткового мозку незрілих клітин, які після короткочасного первинного перебування в крові надходять у тканини. Звідси вони знову можуть повертатися у кровотік, де циркулюють тепер вже протягом багатьох діб, створюючи ефект еозинофілії.
4. Лімфоцити – на відміну від більшості інших клітин крові, зберегли здатність до проліферації і диференціюванні. Утворюються лімфоцити в кістковому мозку, виходять в кровотік. Але більшість з них мають повторно дозріти у лімфоїдних органах.
Розрізняють первинні лімфоїдні органи (тимус, кістковий мозок) і вторинні лімфоїдні органи (лімфатичні вузли, селезінка, лімфоїдна тканина ШКТ). У ці утворення лімфоцити надходять із кісткового мозку і тимусу (Т і В -лімфоцити). У вторинних ЛО відбувається проліферація лімфоцитів у відповідь на надходження у організм чужорідного білка-антигена. Вказані Т- л/ц здатні да рециркуляції: вони знову з’являються в крові і надходять до нових ділянок вторинних лімфоїдних органів, де утворюють колонії, що синтезують ці антитіла.
Т-лімфоцити. Формують тканинний імунітет(протипухлинний). Їх функції:
а) Імунний контролер – вступає в контакт з АГ, запам’ятовую його антигенну структуру і визначає програму біосинтезу АТ (імуноглобулінв). Синтез АТ виконує В-система.
б) синтез речовин, які активують фагоцитоз і захисні запальні реакції.
в) активують регенерацію, знешкоджують загиблі, старі клітини, пухлини.
г) регулюють кровотворні функціі – знешкоджують чужорідні стовбурові клітини кісткового мозку.
Т-лімфоцити поділяються на кілери, хелпери (помічники В-системи), супресори (пригнічують В-систему).
В-лімфоцити формують кров’яний імунітет. З них утворюються плазматичні клітини – фабрика антитіл. Антитіла з’єднуються з антигеном, утворюється імунний комплекс АГ-АТ, який виводиться через нирки.
5. Моноцити – клітини, які ще зберігають потенцію до подальшого розвитку. Моноцити циркулюють в крові від 30 хв до 72 годин, потім потрапляють в тканини і перетворюються на макрофаги. Макрофаги можуть трансформуватися в інші клітини (остеокласти).
Функції:
а) фагоцитоз бактерій, старих і ушкоджений клітин
б) синтезують деякі компоненти комплементу, інтерферону.
в)постачають у кровотік ендогенний піроген (білок, що діє на центр терморегуляції)
г) беруть участь у формуванні АТ, захист від пухлин, відторгнення трансплантату (функції тканинного імунітету).
Фагоцитоз –це поглинання чужорідних об’єктів. Здатністю до фагоцитозу володіють мікро- і макрофаги. Макрофаги утворюють моноцитарно-фагоцитарну систему фагоцитів. Ця система розміщена у різних органах і тканинах: у сполучній тканині, навколо базальних мембран кровоносних судин, у легенях (альвеолярні макрофаги), в печінці (клітини Купфера). Макрофаги здатні до міграції та цілеспрямованого хемотаксису. Речовини, що визначають напрямок руху макрофагів називають хемоантрактантами (лімфокіни, продукти деградації фібрину, колагену, клітин тощо). Поступово підключення різних хемоантрактантів забезпечує постійний приплив нових макрофагів із судинного русла. Велике значення для знешкодження чужорідного агента мають фактори, які гальмують міграцію макрофагів і затримують їх у вогнищі запалення: інтерферон, імунні комплекси, гепарин, глюкокортикоїди. Підсилюють міграцію макрофагів нуклеїнат натрію, левамізол.
Стадії фагоцитозу.
Наближення фагоцита і мікроба в результаті позитивного хемотаксису.
Адгезія мікроорганізму на поверхні фагоцита.
Утворення псевдоподій та поглинання мікроорганізму.
Внутрішньоклітинне перетравлення чужерідної частинки і виділення продуктів розпаду за межі клітини.
Імунна система – захист організму від чужерідних речовин живої і неживої природи. Вона сформована трьома системами: А-система (моноцити, перші контактують с антигеном), Т – система (протипухлинний імунітет), В – система (виробляе плазмоциди, антитіла). Антитіла бувають неспецифічні (до будь-якого антигену) і специфічні (до визначеного антигену). АТ (специфічне) підходить до АГ як ключ до замку. Утворюється імунний комплекс АГ-АТ. Він виводиться з організму через нирки.
ІІ. Тромбоцити, або кров’яні пластинки – без’ядерні бліді формені елементи крові, що мають розмір 2-3 мкм. Це фрагменти клітин – мегакаріоциту, яка в кістковому мозку розпадається на тисячі овальних частинок – кров’яних пластинок. Кількість Тц: 180-320 х 10 9 в 1 л. Тромбоцити циркулюють у крові і частково знаходяться у депо (кістковий мозок, селезінка, легені, печінка). Звідси при потребі вони виходять у кровоносне русло. Кількість тромбоцитів збільшується під час приймання їжі, м’язової роботи. Тц в крові циркулюють протягом 8-12 діб. Потім вони руйнуються у селезінці, печінці, легенях або прилипають до ендотелію кровоносних судин (там вони виконують ангіотрофічну функцію – стінка капілярів стає міцнішою).
Функції Тц:
І. Захисна - участь у згортанні крові (гемокоагуляція) і припиненні кровотечі завдяки наступним їх властивостям:
1) На мембрані і в гранулах цитоплазми Тц містять БАВ, які призводять до згортання: серотонін, адреналін, кальцій, фібриноген, ректрактозим, попередник тромбопластину (транспортна функція Тц).
2) Властивості Тц - здатні прилипати (адгезія) до ушкодженої стінки судин, скупчуватися і склеюватися (агрегація, аглютинація), формуючи таким чином згусток крові.
ІІ. Ангіотрофічна функція - живленні Тц ендотелію судин.
При тромбопенії (зменшенні кількості тромбоцитів) ця функція порушується, тому еритроцити виходять через ендотеліальні клітини судин і утворюються крововиливи.
ІІІ. Гемостаз.
Система гемостазу забезпечує збереження крові в рідкому стані, запобігає кровотечі і відповідає за структуру стінок кровоносних судин. У разі ушкодження судин завдяки цієї системи швидко зупиняється кровотеча. Система гемостазу бере участь в підтриманні об’єму крові на оптимальному рівні. Гемостаз здійснюється завдяки взаємодії стінок кровоносних судин, формених елементів крові, факторів (40), що містяться в плазмі крові. Фактори плазми поділяються на дві групи: ті, що забезпечують процес зсідання крові (коагулянти), і ті, що руйнують (антикоагулянти).
Згортальна система крові – це захисна реакція організму, яка попереджує значну втрату крові. Кров, яка вийшла із кровоносного русла, згортається протягом 3-4 хв і з рідкого стану переходить у драглистий згусток, закриваючи таким чином місце пошкодження судин.
Кровотеча зупиняється в два етапи :
Судинно-тромбоцитарний гемостаз (первинний) – рефлекторний спазм судин, адгезія тромбоцитів, агрегація (скупчення) тромбоцитів, ущільнення згустка крові (ретракція). Через кілька хвилин кровотеча з невеликих ран зупиняється. Цей гемостаз достатній для маленьких судин.
Тромбоцитарний гемостаз (коагуляційний, вторинний) – формування справжнього тромбу (з ниток фібрину). Цей етап йде одночасно з первинним гемостазом. Кров’яний згусток не витримує тиску крові (витискується з пошкодження), тому і включається вторинний гемостаз. Особливо це має значення у середніх і крупних судинах.
Згортання – складний ферментативний процес, в якому беруть участь не тільки тромбоцити і й так звані фактори згортання плазми.
Факторів згортання плазми нараховують тринадцять і їх позначають римськими цифрами.
І – фібриноген, ІІ- протромбін, ІІІ-тканинний тромбопластин, ІУ – іони кальцію,
У- Ас-глобулін (протромбіназа), УІ –проакцелерин, УІІ –проконвертив, УІІІ – антигемофільний глобулін, ІХ –плазматичний компонент тромбопластину, фактор Кристмаса, Х – фактор Стюарта-Прауера, протромбокіназа, ХІ – плазменний попередник тромбопластину, ХІІ – фактор Хагемана, ХІІІ – фібринстимабілізуючий фактор, фібриназа. Усі ці фактори поділяються на дві групи : ферменти (їх попередники) фактори ХІІ,ХІ,Х,ІХ,УІІ,ІІ; неферменти – фактори І,ІУ,У,УІІІ (І – білок, інші – активатори ферментативних процесів).
Стадії згортання крові:
І стадія –формування фермента проттромбокінази ( в його основі – ліпідний фактор), звільненого з мембран пошкоджених тканин, судин, тромбоцитів і еритроцитів. Для цього необхідні іони кальцію, фактори згортання. Процес триває 5- 7 хв.
ІІ стадія – утворюється із білка протромбіна фермент тромбін (за участю протромбокінази, віт К, іонів Са) за 2-5 с. Вітамін К необхідний для утворення протромбіна в печінці.
ІІІ стадія – розчинений білок крові фібриноген перетворюється в нерозчинний фібрин (під дією тромбіну). Для цього необхідні іони Са і фактори згортання. Фібрин – це нитки, які утворюють сітку. Під дією тромбостеніну, який виходить із пошкоджених еритроцитів, відбувається рефракція (стиснення) згустку крові, яка завершується через 2- 3 години (згусток не пропускає навіть сироватку).
Протизгортальна система (фібринолітична).
До антикоагулянтної системи належать антикоагулянти (антитромбін ІІІ, гепарин). Гладка поверхня судин теж сповільнює згортання крові. В крові існує також система, яка може розчинити фібрин, що утворюється під час згортання. До неї належить плазмін, а процес розщеплення фібрину називається фібриноліз.
Більшість людей помирає від захворювань, що пов’язані із підвищенням згортання крові. Підвищення згортання крові називається гіперкоагуляція, а зниження – гіпокоагуляція. Гіперкоагуляція супроводжується прискореним утворенням тромбів, а гіпокоагуляція – кровотечею. Гіперкоагуляція спостерігається при стресових ситуаціях, що пов’язано з надмірним виділенням адреналіну.
Лекція №12.
Тема: Травлення в кишці. Роль печінки і підшлункової залози в травленні.
План:
І. Травлення в 12-палій кишці. Секрет підшлункової залози. Регуляція панкреатичної секреції.
ІІ. Роль печінки в травленні. Жовч – склад. Утворення, властивості. Види жовчі, методи дослідження. Регуляція утворення жовчі.
ІІІ. Кишковий сік. Роль в травленні, регуляція. Моторна діяльність тонкої кишки, регуляція.
ІУ. Травлення в товстій кишці. Роль мікрофлори. Моторика товстої кишки, регуляція. Акт дефекації. Всмоктування різних речовин, регуляція.
І. Кишковий відділ травного каналу складається з тонких і товстих кишок, які функціонують як єдина система, тому що гідроліз хімусу тут відбувається у лужному середовищі, яке формується на початку процесу. У кишках відбувається остаточне розщеплення всіх полімерів до мономерів, їхнє всмоктування і формування калових мас з неперетравлених залишків їжі.
Тонкі кишки. Особливе функціональне значення має 12-пала кишка. Харчовий хімус у 12-палій кишці перетравлюється під дією травних соків підшлункової залози, а також жовчі. Крім того, в стінці кишки розташовані невеликі залози (бруннерові), сік яких містить протеолітичні (подібні до пепсину), амілолітичні і ліполітичні ферменти. В результаті білки, жири і вуглеводи підлягають розщепленню до речовин, які надалі можуть всмоктуватися в кров і лімфу. Сік дванадцятипалої кишки нейтралізує кислий хімус. Регуляція соковиділення: нервова (блукаючий нерв) і гуморальна (секретин). Пригнічує секрецію симпатична нервова система (норадреналін). Тому при збудженні СНС (стреси) можливе утворення виразок 12-палої кишки.
Підшлунковий сік – безколірна рідина, має лужну реакцію, рН=8-8,5 (створюється гідрокарбонатами), має велику кількість ферментів в повному складі. За добу виробляється 1,5-2,0 л соку.
Ферменти:
Протеолітичні ферменти – трипсин, хемотрипсин, карбоксиполіпептидаза розщеплюють білки до пептидів і амінокислот. Ці ферменти виробляються в неактивному стані. Трипсиноген переходить в трипсин під впливом ентерокінази, яка утворюється в 12-палій кишці. Трипсин активує хемотрипсиноген, перетворюючи його в хемотрипсин. Протеолітичні ферменти діють тільки в лужному середовищі.
Амілолітичні ферменти – амілаза, мальтаза, лактаза гідролізують вуглеводи. Амілаза розщеплює полісахарид крохмаль до дисахаридів. Мальтоза розщеплює дисахарид мальтозу до глюкози. Лактаза розщеплює молочний цукор до моносахариди глюкози.
Ліполітичні ферменти – ліпаза, фосфоліпаза розщеплюють жири і фосфоліпіди до жирних кислот і гліцерину. Ці ферменти розщеплюють жири, які раніше були емульсовані жовчю.
Регуляція секреції підшлункового соку:
Нервова - секреція соку починається з умовних подразників, ще до потрапляння їжі в рот (запах, вигляд страви).Посилюється секреція при дії їжі на рецептори порожнини рота. Секреторний нерв – блукаючий. Центр соковідділення – в довгастому мозку. Симпатичні нерви гальмують виділення соку (негативні емоції, біль).
Гуморальна – при потраплянні хімусу в 12-палу кишку. Це гормони – секретин, холецистокінін-панкреазимін (виробляються слизовою 12-палої кишки). Вони стимулюють соковидділення. Ці гормони утворюються тільки при потраплянні кислого хімусу в лужне середовище кишки. У людей з порушенням секреції соляної кислоти не утворюються ці гормони, що призводить до різкого зменшення панкреатичного соковідділення.
Фази секреції підшлункового соку:
Мозкова фаза працюють умовно-рефлекторні і бузумовні механізми. Еферентний шлях передає сигнали від центра довгастого мозку по блукаючому нерву. Секреція починається через 1-2 хв після початку прийому їжі. Виділяється помірна кількість соку.
Шлункова фаза – рефлекторне виділення соку за рахунок подразнення хімусом рецепторів шлунку, тут також приєднуються гуморальні фактори.
Кишкова фаза – при потраплянні хімусу в 12-палу кишку. Утворюється два основних ГІГ – секретин і холецистокінін-панкреозимін. Вони посилюють виділення соку. Також секрецію посилюють гастрин, серотонін, бромбезин, субстанція Р, інсулін. Гальмує виділення соку глюкагон, кальцитонін, соматостатин.
ІІ. Роль печінки в травленні. Жовч – склад. Утворення, властивості. Види жовчі, методи дослідження. Регуляція утворення жовчі.
Печінка виконує ряд таких важливих функцій:
Травна – утворення жовчі. Жовч утворюється гепатоцитами постійно (800 -1000 мл за добу) і потрапляє в 12-палу кишку. Вона має лужну реакцію, нейтралізує кислий хімус, що надійшов зі шлунка.
Біологічний фільтр (бар’єр) – для крові, яка притікає з органів травлення. Інактивуються токсини, які потрапили з їжею або утворилися в кишечнику (індол, скатол, фенол), інфекційні та токсичні агенти, деякі лікувальні речовини.
Захисна – печінка бере участь в утворенні антитіл і факторів загортальної й протизгортальної системи крові.
Обмін гормонів і вітамінів. В неї відбувається інактивація стероїдних гормонів, інсуліна, глюкагона, вазопресину, гормонів щитоподібної залози. В печінці синтезується вітамін А. Для всмоктування жиророзчинних вітамінів необхідна жовч.
Обмін білків, жирів, вуглеводів. В печінки утворюються білки плазми крові, сечовина, тригли цериди, фосфоліпіди, холестерин, глікоген (запаси глікогену до 20% маси органа).
Орган кровотворення в пренатальному періоді життя. В постнатальному періоді тут депонує антианемічний фактор. Тут синтезуються регулятори кровотворення (еритропоетин, тромбоцитпоетин). Тут руйнуються еритроцити і дезінтоксикуються продукти руйнування Нв.
Вироблення тепла при обміні речовин.
Екскреторна функція – виведення з організму більш 40 сполук, які синтезуються в самої печінці або є метаболітами крові (холестерин, жовчні кислоти, білірубін, сечовина, спирти).
Жовч накопичується в жовчному міхурі, звідки надходить у 12-палу кишку тільки в процесі травлення при наявності в ній жирної їжі. Поза процесом травлення сфінктер Одді (в зоні загальної жовчної протоки) закритий і жовч потрапляє в міхур. Концентраційна здатність жовчного міхура, об’ємом 50-80 мл, дозволяє збирати жовч, яка постійно утворюється в печінці, навіть в перебігу 12 годин (до 400-450 мл печінкової жовчі). Існує печінкова і міхурові жовч. Міхурові жовч, за рахунок всмоктування води стінкою міхура, густіша і має темніше забарвлення.
В кишечнику жовч виконує слідуючи функції:
1) Емульгація жирів – роздрібнює для гідролізу їх ліпазами.
2) Утворює комплекси з жирними кислотами, забезпечуючи їх всмоктування. Без жовчі всмоктується лише 60% жиру, який потрапляє з їжею.
3) Підвищує активність панкреатичних і кишечник ферментів.
4) Регулює процеси жовчоутворення.
5) Забезпечує бактеріостатичний ефект.
6) Солі жовчних кислот стимулюють моторику кишечнику.
7) Виводить деяких продуктів обміну – жовчні пігменти, холестерин.
Склад жовчі:
Жовчні кислоти –в гепатоцитах із холестерина утворюються жовчні кислоти (холева і хенодезоксихолева). В жовчі вони з’єднуються з глікоколом і таурином. Зазвичай печінкова жовч вміщує 75% глікохолевих і 25% таурохолевих кислот (перші збільшуються при вуглеводних дієтах, другі – при високобілкових). Жовчні кислоти забезпечують процеси засвоєння жиру. З калом виводиться 10-15% ЖК, а інші ЖК активно ре абсорбуються і повертаються через ворітну вену в печінку.
Жовчні пігменти (білірубін, білівердин). Вони є кінцевими продуктами розпаду гемоглобіну. Нерозчинний у воді білірубін переноситься до печінці у зв’язку з альбуміном. Він має червоно-жовтий колір, і надає характерне забарвлення жовчі. Зеленого кольору білівердин в жовчі складає невелику кількість. З жовчю в кишечник за добу виділяється 200-300 мг білірубіну, основна частина якого виділяється з калом, забезпечує його колір (при порушенні потрапляння жовчі в кишечник кал стає білим).
Вода, електроліти, холестерин, лецитин. Холестерин ( жир крові), утворюється в печінці 1,5-2,5 г за добу (0,5 г потрапляє з їжею), він з’єднується з білками (ліпопротеїди), далі виводиться з організму. Лецитин емульгує холестерин і триглицериди, допомагає утримувати холестерин в розчиненому стані, щоб він не відкладався на стінки судин.
Регуляція секреції і виділення жовчі.
1. Нервова – на основі умовних і безумовних рефлексів, зв’язаних з прийняттям їжі (імпульси йдуть в довгастий мозок, до ядер блукаючого нерва, потім по блукаючому нерву імпульси йдуть до м’язів жовчного міхура). Виділяється невелика кількість жовчі. Період триває 7-10 хвилин. Потім починається більш активні скорочення і розслаблення жовчного міхура, що при відкритому сфінктері Одді призводить до виділенню міхурової жовчі в кишечник. Після опорожнення жовчного міхура в кишечник потрапляє менш концентрована жовч прямо з печінки.
2. Гуморальна – під час травлення вдвічі збільшується жовчовиділення. Скорочення жовчного міхура і жовчних протоків при одночасному розслабленню сфінктерів викликає холецистокінін– панкреазимін (виробляється слизовою дванадцятипалої кишки). Секреція багатої бікарбонатами жовчі стимулюється секретином. Посилюють жовчовиділення жири, яєчний жовток, сульфат магнію, сама жовч.
ІІІ. Кишковий сік. Роль в травленні, регуляція. Моторна діяльність тонкої кишки, регуляція.
В тонкій кишці під впливом кишкового соку відбувається остаточний гідроліз усіх харчових речовин до стадії мономерів і всмоктування їх в кров і лімфу. Травлення в тонкій кишці відбувається спочатку в її порожнині – порожнинне травлення, а потім в зоні кишкового епітелію за допомогою ферментів, які фіксуються на мікроворсинках кишкового епітелію – пристінкове або мембранне травлення.
Кишкові залози знаходяться в слизовій оболонці всієї тонкої кишки. Кишечник сік – каламутна, в’язка рідина. Кишечник сік складається з 20 ферментів.
Склад кишечного соку:
Пептидази (протеолітичні ферменти)
Ліпаза (ліполітичні ферменти)
Амілаза, мальтаза, лактаза (амілолітичні ферменти).
Неорганічні речовини - бікарбонати, хлориди, фосфат натрію, кальцій, калій.
Органічні речовини – білки, амінокислоти, слиз (захист слизової оболонки).
Регуляція:
Рефлекторна – сік виділяється тільки при потраплянні хімусу в кишечник (подразнення механо- і хеморецепторів слизової). Секреторний нерв – блукаючий.
Гуморальна – секретин, холецистокінін-панкреазимін, мотилін.
Моторна діяльність тонкої кишки:
Перистальтика – при узгодженому скороченні циркулярних і поздовжніх м’язів стінки кишки. При цьому вище харчового хімусу утворюється звуження, а нижче – розширення порожнини кишки. Харчова кашка витискується в розширену частину кишечника. Хвилеподібні рухи проштовхують харчову кашку до прямої кишки. Пропульсивна перистальтика (потужні хвилі вздовж всього тонкого кишечнику) проштовхує залишки хімусу в товсту кишку наприкінці процесу травлення (через 3,5-4 години після прийому їжі) і триває 4-5 годин. Отже, через 8-10 годин після прийому їжі залишки хімусу потрапляють повністю в товстий кишечник.
Ритмічна сегментація – періодичне скорочення циркулярного шару м’язів на ділянках шириною 1,0-1,5 см, на відстані один від одного 15-20 см.
Маятникоподібні рухи – скорочення невеликої ділянки поздовжніх м’язів, завдяки чому стінка зміщується по відношенню до хімусу. При цьому хімус переміщається «вперед-назад» і тим самим ретельно перемішується.
Регуляція моторики:
Автоматія – завдяки пейсмекерам тонкої кишки (де відбувається спонтанна деполяризація). «Вузли» автоматії знаходяться при впадінні загальної жовчної протоки в 12-палу кишку і в клубовій кишці.
Рефлекторна регуляція (місцева)– забезпечують нервові сплетення кишки. Подразнення виникають при розтягненні стінки кишки хімусом. Вона регулюється вегетативною системою: ПСНС – збуджують, а СНС – гальмує скорочення.
Гуморальна – серотонін, вазопресин, окситоцин, гістамін, гастрин, мотилін. ХЦК-ПЗ, брадикінін, кислоти, луги. Всі вони посилюють моторику.
ІУ. Травлення в товстій кишці. Роль мікрофлори. Моторика товстої кишки, регуляція. Акт дефекації. Всмоктування різних речовин, регуляція.
Залишки їжі, яка не перетравилася в тонких кишках, потрапляє через ілеоцекальну заслонку в сліпу кишку ( за добу 300-500 мл).В товстих кишках відбувається концентрація хімусу за рахунок всмоктування води. Також продовжується всмоктування електролітів, водорозчинних вітамінів, жирних кислот, вуглеводів. При відсутності механічних подразників (при відсутності хімусу) виділяється незначна кількість соку. При подразненні хімусом, сікоутворення збільшується в 8-10 разів. Сік вміщує незначно ферментів, слиз. Перетравлюючи здатність соку незначна. Основна функція соку – захист слизової оболонки від механічних, хімічних пошкоджень і забезпечення слабко лужної реакції (8,0).
Мікрофлора товстої кишки: 90% безспорові анаероби, 10% - молочнокислі бактерії, кишкова паличка, стрептококи й спороносні анаероби.
Функції:
1)кінцевий розклад залишків неперетравлених речовин і компонентів травних секретів
2) синтез вітамінів групи В і К, різних БАР (ГАМК, глутамат)
3) участь в обміні речовин
4) імунний бар’єр через пригнічення патогенних мікробів
5) стимулює розвиток імунної системи організму
Під дією мікробів неперетравлені вуглеводи зброджуються до молочної і оцтової кислот, алкоголю, СО2, Н2О. З білками відбувається гнилісний розклад і утворюються токсичні з’єднання і гази: індол, скатол, водень, сернистий газ, метан. Ці токсини йдуть в печінки, там вони знешкоджуються. Балансоване харчування зрівноважує процеси гниття і бродіння (кислі продукти бродіння перешкоджають гниттю).
Регуляція секреції соку в товстих кишках відбувається як нервовим шляхом (через механо- і хеморецептори), так і гуморальним (секретин, мотилин, ХПК-ПЗ). В товстій кишці секреція також визначається місцевими рефлексами, які обумовлені механічним подразненням хімусу.
Моторна функція товстої кишки. Товста кишка виконує функцію депо неперетравлених залишків хімусу і формує калові маси. Проходження хімусу відбувається повільно (за 15-2 діб). Це сприяє тривалий контакт зі стінкою кишки, створює благо приємні умови для закінчення всмоктування, здебільше води і солей.
Зовнішній повздовжній шар м’язів розташований у вигляді смуг, і знаходиться в постійному тонусі. При цьому скорочення окремих ділянок циркулярного м’язового шару утворює складки і здуття (гаустри). Хвилі гаустрації повільно прокативаються по товстій кишці. Проштовхування їжі виникає в результаті пропульсивної перистальтики, 3-4 рази за добу.
Регуляція моторной функції:
Нервова – через місцеві рефлекси (інтрамуральні нервові сплетення). Механічні і хімічні подразники стимулюють перистальтику. Вищерозташовані відділи ВНС корегують місцеві рефлекси: парасимпатика стимулює, а сим патика пригнічує перистальтику.
Гуморальна – серотонін збуджує. А адреналін гальмує моторику товстої кишки.
Дефекація. За добу утворюється 100-200 г кала.
Склад:
1. Вода – 75-80%
2. Сухий залишок – целюлоза та інші неперетравлені речовини, бактерії 10-30 %, 10-15% неорганічних речовин, 5% жиру, злущеного епітелію, слизу.
Колір – результат розкладу пігментів жовчі. Запах обумовлений сірководнем, індолом, скатолом.
Механізм дефекації:
Подразнення рецепторів прямої кишки.
Аферентні нерви – статевий, тазовий нерв. Імпульс йде до крижового відділу спинного мозку (центр дефекації).
Еферентні імпульси йдуть по парасимпатичному нерву до внутрішнього анального сфінктеру. Розслаблення внутрішнього сфінктеру запускає рефлекси посилення перистальтики прямої кишки, розслаблення зовнішнього сфінктеру, скорочення м’язів черевного пресу і діафрагми. Участь скелетних м’язів забезпечує можливість свідомого впливу на процеси дефекації.
Рефлекс дефекації оказує ряд рефлекторних впливів на різні системи органів. Так, серцеві скорочення учащаються на 20 ударів за хвилину. Можливо збільшення систолічного тиску до 60 мм рт.ст., діастолічного – до 20 мм рт.ст.
Всмоктувальна функція ШКТ. Всмоктування – це переніс речовин із порожнини кишечнику в кров і лімфу.
Сприяє всмоктуванню:
1.Скорочення ворсинок – йде стискування капілярів і відтоку крові і лімфи від ворсинок. Розслаблення ворсинок утворює присмокту вальну дію – поліпшує всмоктування.
2. Перистальтика кишок –підвищення внутрішньо порожнистого тиску, забезпечує приріст фільтраційного тиску.
3. Посилення кровозабеспечення слизової кишки в 2-3 рази. Кров від шлунка і кишок проходить через печінку, а потім йде в загальний кровоток.
Від слизової рота, прямої кишки, кров одразу йде в загальний кровоток, минаючі печінку.
Механізми всмоктування: пасивний осмос, дифузія, активний енергозалежний транспорт.
Лекція № 16.
Тема: «Фізіологія системи виділення. Регуляція сечоутворення і сечовиділення»
План:
Система виділення. ЇЇ будова, функції.
Органи виділення( нирки, шкіра, легені, травний канал), їх участь в підтриманні гомеостазу.
Нефрон – функціональна і структурна одиниця нирки. Кровообіг в нирках.
Основні процеси сечоутворення: фільтрація, реабсорбція, секреція.
Кінцева сеча. Коефіцієнт очищення (кліренс).
Регуляція сечоутворення. Роль ренін-ангіотензин-альдостеронової системи в регуляції водно-сольового обміну.
Роль нирок у регуляції кислотно – основного стану внутрішнього середовища.
Сечовиділення. Його регуляція.
І. Система виділення. В процесі життєдіяльності в організмі людини утворюються значна кількість продуктів розпаду органічних речовин (метаболітів), частина яких не використовується клітинами та може бути шкідливою для організму. Метаболіти, які утворюються в результаті обміну речовин та виводяться із організму звуть екскретами, а органи, які сприяють їх виведенню екскреторними (видільними). До органів виділення із організму метаболітів , чужорідних і токсичних речовин відносяться нирки, сальні та потові залози шкіри, легень, молочні залози в період лактації і травний тракт. Видільна функція шкіри забезпечується потовими та сальними залозами. З потом виділяється не тільки вода і солі, але й сечовина, азот, токсичні речовини (ацетон, жовчні кислоти). Концентрація сечовини в поті може бути в 2 рази вища, чим у плазмі крові, молочної кислоти – в 4 рази, калію – в 1,2 рази. З потом виводяться також іони натрію, хлору і т.п. Слинні залози можуть виводити сечовину, з’єднання ртуті, вісмуту, брому, йоду. Слизова оболонка шлунку здатна виводити сечовину, з’єднання ртуті, миш’яку, хлороформу, брому, саліцилати. Тонкий кишечник виводить сечовину, сечову кислоту, іони калію, кальцію, фосфору, хлору, з’єднання ртуті, нікелю. Підшлункова залоза виводить сечовину, сечову кислоту, солі кальцію, цинку, залізу, ртуті, срібла, брому. Печінка виводить сечовину, глютамін, креатинін, холестерин, жовчні пігменти. Через легені виводяться СО2, ефір, хлороформ, ацетон, алкоголь. Шкіра, органи ШКТ, легені виводять також значну кількість води. Значення екстраренальних шляхів виведення різко підвищується при ураженні нирок. Хоча повністю замінити їх вони не в змозі.
ІІ. Основним органом екскреції є нирки. Процес утворення і виділення сечі із організму звуть діурезом. Функціональна активність нирок пов’язана з їх будовою. Нирки розташовані на обох боках поперекового відділу хребта, вкриті сполучною тканиною і мають масу 200-250 грам. На фронтальному розрізі є два шари: корковий (темно-червоний) та піраміди(7-10) або мозковий (світліший).
Кровопостачання нирок. Нирки получають кров із ниркової артерії. Артерія в нирці поділяється на велику кількість дрібних судин – артеріол, які приносять кров до клубочка (приносна артеріола), які потім розпадаються на капіляри (перша сітка капілярів). Капіляри судинного клубочка, зливаючись, утворюють виносну артеріолу, діаметр якої в 2 рази менша діаметру приносної. Виносна артеріола знов розпадається на сітку капілярів, які обплітають канальні (друга сітка капілярів). Таким чином, для нирки характерно наявність двох сіток капілярів: капіляри судинного клубочка й капіляри, які обплітають канальні.
Тиск крові в капілярах судинного клубочка вищий, ніж у всіх капілярах тіла. Він дорівнює 70-90 мм рт.ст., що складає 60-70% від величини тиску в аорті. В капілярах, які обплітають канальні нирки, тиск невеликий – 20-40 мм рт.ст.
Через нирки вся кров проходить за 5 хвилин. Такий кровоток дозволяє повністю виводити усі непотрібні речовини. Артеріальні капіляри переходять в венозні, які в подальшому зливаються в вени. На мікрорівні нирки складаються із великої кількості нефронів (нирка має біля 1 млн. нефронів). Нефрон – це функціональна одиниця нирки. Нефрон має слідуючи відділи: 1) ниркове (мальпігієво тільце) – це судинний клубочок, капсула Шаумлянського-Боумена, 2)канальні – проксимальний звивистий, петля Генне, дистальний звивистий, 3) збиральна трубочка. Збиральна трубочка відкривається у верхівці сосочка піраміди, який виступає в порожнину ниркової миски. Ниркова миска відкривається в сечовід, який впадає в сечовий міхур.
Функції нирки:
Виведення шлаків – метаболітів азотистого обміну
Виведення чужорідних речовин.
Виведення надлишок органічних і неорганічних речовин
Підтримка постійності осмотиску крові.
Підтримка іонного балансу організму.
Підтримка кислото-основного стану
Участь в обміні білків, жирів, вуглеводів.
Участь в регуляції кровообігу.
Участь в регуляції об’єму циркулюючої крові.
Секреція БАР і ферментів (брадикініни, простагландіни, урокінази)
Інкреторні - регуляція еритропоєзу (еритропоетін), підвищення АТ (ренін), участь в фібринолізі (урокіназа), участь в реабсорбції кальцію в нирках.
ІІІ. Основні процеси сечоутворення: фільтрація, реабсорбція, секреція.
У всіх структурах нефрону відбувається процес сечоутворення, завдяки чому із крові утворюється кінцева сеча.
1. Клубочкова фільтрація (утворення первинної сечі) – в капілярах клубочків відбувається фільтрація із плазми крові води з розчиненими у неї неорганічними і органічними речовинами. Фільтраційна ниркова мембрана складається із трьох шарів: ендотелія капілярів, базальної мембрани і епітеліальних клітин – подоцитів. Всі вони мають вікна, через які проходить вода і речовини плазми, але обмежують проходження білків (особливо обмежує базальна мембрана). Також проходження речовин залежить від заряду – білки і стінки пор мають негативний заряд. В нормі в фільтраті (первинної сечі) може визначатися майже всі речовини, які є в плазмі крові, за виключенням білків і клітин крові. Фільтрація відбувається без затрати енергії і є типовим пасивним транспортом речовин. Об’єм фільтрату та швидкість його утворення залежить від ефективного фільтраційного тиску (ЕФТ). ЕФТ є результатом дії сил, які фільтрують вміст крові і протидіють фільтрації. Так, фільтруюча сила є тиском крові в капілярах клубочка (Рк=70 мм рт. ст..). Протидіючою силою фільтрації є онкотичний тиск білків плазми крові (Ро=30 мм рт.ст.) і гідростатичний тиск фільтрату в капсулі клубочка (Рф=20 мм рт.ст.). ЕФТ = 70-(30+20)=20 мм рт.ст. В фільтрат потрапляє десь приблизно 1/5 частини плазми, яка проходить через нирки. В результаті, за добу утворюється 150-180 л фільтрату (вся плазма крові очищується за добу нирками 60 разів). Склад первинної сечі подібний до плазми крові, але має мало білків.
2. Канальцієва реабсорбція (утворення вторинної сечі). Зворотне всмоктування різних речовин із первинної сечі у кров називається реасорбцією і відбувається вона в канальцях нефрону. Це стосується води, глюкози, амінокислот, солей. Завдяки реабсорбції залишається 1% від первинної сечі – 1,5 л вторинної сечі.
Механізми реабсорбції:
Пасивний транспорт – відбувається за електрохімічним, концентраційним і осмотичним градієнтом. Таким чином потрапляє назад у кров вода, вуглекислий газ, деякі іони, невелика кількість сечовини.
Активний транспорт – проходить проти електрохімічного і концентраційного градієнтів. Для його здійснення необхідна енергія АТФ та спеціальні переносники і обов’язково приєднані до них іони натрію. Активно транспортуються глюкоза, амінокислоти, натрій.
Реабсорбція відбувається в різних відділах нефрону неоднаково:
Проксимальні канальні – всмоктується глюкоза, натрій, калій, вода. Це обов’язкова реабсорбція.
Петля Генле – всмоктується вода і іони натрію за рахунок поворотно-протипотокової системи (обов’язкова реабсорбція – з енергією АТФ). Проходячи через низхідний відділ петлі Генле, сеча віддає воду, згущується, стає більш концентрованою. Віддача води відбувається пасивно за рахунок того, що в висхідному коліні відбувається активна реабсорбція іонів натрію. Потрапляючи в тканинну рідину, іони натрію підвищують у неї осмотичний тиск і тим самим сприяють притягненню в тканинну рідину води з низхідного колена. В свою чергу підвищення концентрації сечі в петлі Генне за рахунок реабсорбції полегшує перехід іонів натрію із сечі в тканинну рідину. Отже, в петлі Генле відбувається реабсорбція великої кількості води і іонів натрію.

Дистальні канальні - подальше всмоктування іонів натрію, калію, води та інших речовин, яке залежить від їх концентрації в крові (факультативна реабсорбція). Отже, дистальні канальні підтримують постійність концентрацій іонів натрію і калію в організмі.
Збірні трубочки – подальше всмоктування води. Збірні трубочки проходять через мозковий шар нирок, в якому тканинна рідина має високий осмотичний тиск і тому притягує до себе воду.
3. Канальцієва секреція. Ряд сполук , які повинні виводитися з сечею, можуть абсорбуватися і секретуватися. Секреція – це процес, який направлений на активний перехід речовини із крові через клітини канальцівв їх просвіт, в первинну сечу. Вона може бути активною і пасивною.
Секретуються такі речовини:
А) сечовина – може секретуватися в петлю Генне.Б) Слабкі органічні кислоти і луги (парааміногіпурова кислота)В) Аміак – принцип неіонної дифузії лежить в основі виведення аміаку, який має високу розчинність в жирах і легко приникає через мембрану в сечу. Але якщо в сечі його не зв’язати, то він також легко може повернутися в клітину або позаклітинну рідину. В сечі перебігає реакція зв’язування Н+ з аміаком, завдяки чому аміак знаходиться у рівновазі з амонієм. Іон амонію погано проникає через мембрану, зв’язуючись з катіонами, виділяється з сечею. Таким чином, кисла сеча (багато Н=) сприяє більший екскреції аміаку.
ІУ. За добу виробляється біла 1,5-2 л вторинної сечі. Інтенсивність сечоутворення залежить від прийнятої рідини, від білкової їжі (збільшується) , від часу доби. Денний діурез інтенсивніший від нічного , оскільки вночі у зв’язку із пониженням АТ знижується фільтраційний тиск, а значить і фільтрація в клубочках нирки. В нормі відношення денного діурезу до нічного визначають за пробою Зимницького і воно становить 2:1. При фізичній роботі зменшується діурез(знижується кровообіг нирок й фільтрація плазми, збільшується потовиділення).
Характеристика вторинної сечі:
Колір – світло-жовтий. При відстою сечі випадає осад із солей і слизу.
Реакція –слабкокисла (4,5-8,0). Сеча стає кислою при вживанні м’яса, а лужною – рослинної їжі.
Відносна густина – щільність сечі 1,018 г/см3 й залежить від прийнятої води (знижується при значної вживанню води). Визначається за пробою Зимниць кого.
Склад – вода, солі (60 г за добу), кінцеві продукти білкового обміну (азот в складі сечовини,сечова кислота, аміак, пурінові луги, креатинін. Можуть бути сліди білка (0,03%). Під час бігу може з’явитися білок, як результат тимчасового збільшення проникності мембрани судинного клубочка нирок. Органічні небілкові з’єднання-солі щавлевої кислоти, молочна кислота, кетонові тіла. Пігменти – урохром і уробілін ( із білірубіна), фарбують сечу в жовтий колір. Гормони – АДГ, кортикостероїди, естрогени, катехоламіни, вітаміни, ферменти.
Фізіологічні методи дослідження функцій нирок.
Найпростіший метод – визначення хімічного складу сечі. Для визначення швидкості виведення різних речовин запроваджено поняття про нирковий кліренс, який відображає швидкість очищення плазми:
Кр=Мр*Кс:Ср.
Кр –кліренс, Мр – концентрація речовини в сечі, Кс –концентрація сечі, яка утворюється за 1хв, Ср –концентрація речовини у крові.
Y. Регуляція сечоутворення. Участь нирок у підтриманні азотистого балансу, параметрів гомеостазу. Роль ренін-ангіотензин-альдостеронової системи в регуляції водно-сольового обміну.
Регуляція сечоутворення:
Нервова – вегетативна нервова система.При дії СНС (подразнення черевного нерва) зменшується сечовиділення(тому що звужуються судини нирок і знижується кількість крові, яка поступає до нирок) і підвищують реабсорбціяю натрію і води. Посилення сечоутворення спостерігається в експерименті при уколі в мозочок, зоровий горб, дно ІУ шлуночка.
Гуморальна:
А) АДГ стимулює реабсорбцію води, адреналін зменшує сечовиділення. Альдостерон збільшує реабсорбцію натрію, сприяє реабсорбції води (зменшення діурезу).
Б) У передсердях при їх перерозтягненні кров’ю утворюється натрій-уретичний гормон (сприяє зменшенню реабсорбції натрію, води, підвищує діурез). Гормони аденогіпофіза та тироксин збільшують сечоутворення.
Ренін-ангіотензинова система організму.
При зниженні АТ, зниженні ниркового кровоточу в нирках утворюється ренін (виробляється в юкстагломерулярному апараті). Під дією реніну (білка) альфа 2 – глобулін плазми перетворюється на ангіотензин І. Потім ангіотензин І трансформується в ангіотензин ІІ (сильний судинозвужувач). Під його впливом підвищується АТ. Він впливає також на наднирникі – підвищується утворення альдостерону (затримує воду в організмі). Вироблення реніну збільшуеться при збуджені СПС. Таким чином, задачею ренін – ангіотензин –альдостеронової системи є підвищення системного АТ і кровотоку через нирки, затримка води і NaCl. При ураженнях нирок, при вагітності(коли знижується нирковий кровоток), ця система активується, що призводить до стійкого підвищення АТ.
YІ. Участь нирок в підтримці КЛС організму обумовлена очищенням крові від нелетучих кислот і луг, які утворюються в процесі обміну, або потрапляють з їжею. Іони, які виводяться в сечу, змінюють її рН від 4,5 до 8,6. При максимальному закисленні сечі іон Н+ виводиться як в вільному стані, так і в зв’язаному стані (з буферними сполуками). Іони водню в сечу потрапляють не тільки з фільтратом. Вони можуть декретуватися в дистальних і проксимальних відділах канальців нефрону. В проксимальних відділах Н+ обмінюється на натрій при реабсорбції останнього в ділянці апікальної поверхні ендотеліальних клітин. При алкалозі рН крові й концентрації в неї НСО№ збільшується, і нирки починають виділяти лужну сечу.
УІІ. Сечовиділення. Його регуляція. Сеча безперервно потрапляє в сечовий міхур (50 мл за годину) й накопичується в сечовому міхурі. При накопиченні тиск збільшується, подразнюються механорецептори. Аферентні імпульси йдуть в спинний мозок до центру сечовипускання (ІІ-ІУ сегменти крижового відділу). Від центру по еферентним шляхам (тазовим нервам) імпульси йдуть до м’яза та сфінктера сечового міхура: м’яз скорочується, несвідомий сфінктер розслаблюється. Одночасно від центра сечовипускання збудження йде в кору головного мозку, де виникає почуття позиву до сечовипускання. Імпульси від кори йдуть до свідомого сфінктеру сечового міхура, виникає акт сечовипускання. Кірковий контроль проявляється в затримці, посиленню або навіть свідомому викликанні сечовиділення. У дітей раннього віку кірковий контроль затримці сечовипускання відсутній, він виробляється з віком.
Лекція №2. Дихальна функція крові.
План:
Еритроцити – будова, кількість, функції. Поняття про еритрон як фізіологічну систему.
Життєвий цикл еритроцитів. Гемоліз. Види.
Осмотична резистентність еритроцитів.
Гемоглобін. Будова. Властивості, Види. Сполуки.
Кількість гемоглобіну. Колірний показник.
Швидкість осідання еритроцитів.
І. Еритроцит – округла, двовгнута клітина без ядра, діаметром близько 7,5 мкм, завтовшки 2,5 мкм. Завдяки такій формі вона має відносно велику поверхню. Оскільки зрілий еритроцит не має ядра, ємкість його збільшується. Відстань від мембрани до найвіддаленішої точки перебування гемоглобіну зменшується. Це забезпечує добрі умови газообміну. Крім того, оскільки еритроцит е без’ядерним і має еластичну мембрану, він легко змінює форму і проходить через найдрібніші капіляри, які діаметром іноді майже у 2 рази менші, ніж еритроцит.
Кількість еритроцитів в нормі у чоловіків 4,5-5,5 х10, у жінок – 3,7- 4,7 в 12 ст. в 1 л. Підвищення кількості еритроцитів зветься еритроцитозом, зменшення – еритропенією (анемія). Еритрон – загальна маса еритроцитів в організмі (в судинах, в депо). Система еритрона – система регуляції еритроцитів:
Функції еритроцитів:
Дихальна – за рахунок Нв.
Захисна – зв’язують токсини (мають на поверхні АТ), беруть участь в згортанні крові.
Ферментативна – носії різних ферментів (холінестерази, метгемоглобін-редуктази).
Підтримує рН крові – через гемоглобінів буфер.
Мембрана еритроцита. МЕ еластична, її товщина близько 10нм, вона у мільйон разів більш проникна для аніонів, ніж для катіонів. Деякі речовини здатні вступати у зв’язок із вбудованими в мембрану молекулами і проходити через неї всередину або транспортуватися на мембрані (це забезпечує транспортну функцію еритроцитів).
.
ІІ. Життєвий цикл еритроцитів. Еритроцити утворюються в червоному кістковому мозку із стовбурової клітини (І клас гемопоезу), клітин – попередників (ІІ, ІІІ клас), еритробластів. Всі вони мають ядра. При дозріванні еритробласта ядро виштовхується через мембрану. Зрілий еритроцит крові – це диференційована без’ядерна клітина, нездатна до подальшої проліферації, здатна циркулювати у кровотоці 100-120 діб, після цього гине. За добу оновлюється близько 1% еритроцитів. Про це свідчить наявність у крові молодих еритроцитів – ретикулоцитів ( від лат. Rete – сітка, основою якої є залишки і-РНК). Після виходу із кісткового мозку у руслі крові вони зберігаються у вигляді ретикулоцитів близько доби. Тому їх концентрація у крові – 0,8 – 1% усіх еритроцитів. Активизація еритропоєзу супроводжується збільшенням числа ретикулоцитів у крові (ретикулоцитоз). Але еритропоез може бути інтенсивнішим не більш ніж у 5-7 разів порівняно з вихідним рівнем. В звичайних умовах за добу утворюється в кістковому мозку близько 25000 еритроцитів на 1мкл крові ( при інтенсивному еритропоезі – до 150000 еритроцитів на 1 мкл). Депо (запасів) еритроцитів в організмі людини немає, тому ліквідація анемії після крововтрат відбувається тільки за рахунок посилення еритропоезу. Збільшення кількості еритроцитів в кістковому мозку відбувається через 3-5 діб, у периферичній крові помітне збільшення кількості еритроцит відбувається ще пізніше. А тому після втрати крові відновлення рівня еритроцитів відбувається не менш як за 2-3 тижня.
Руйнування еритроцитів. Життєвий цикл еритроцитів закінчується їх руйнуванням (гемолізом). При гемолізі Нв виходить із еритроцита.
Гемоліз може бути фізіологічним – при старінні еритроцитів (може відбуватися у руслі крові, в макрофагальній системі: печінка, селезінка), або патологічним – при різних патологіях (укуси змій, уремія, гіпоксії).
Види гемолізу:
Фізіологічний – в судинному руслі йде безперервне руйнування старих еритроцитів. При цьому Нв, що вийшов у плазму, з’єднується з глікопротеїном плазми. Цей комплекс не проникає крізь мембрану нирок, і надходить до печінки, селезінки, кісткового мозку. Тут він розпадається, в печінці перетворюється на білірубін.
Осмотичний гемоліз - відбувається в гіпотонічному розчині.
Механічний гемоліз – при струсах, тривалій ходьбі (марш солдат).
Хімічний гемоліз – кислоти, луги, спирт, ефір визивають денатурацію (згортання) білків і порушення оболонки еритроцитів, з них виходить Нв.
Фізичний гемоліз - - високі температури призводять до денатурації білків оболонок еритроцитів, заморожування крові супроводжується порушенням еритроцитів.
Біологічний гемоліз –при укусах змій, скорпіонів, при малярії, переливаннях несумісної групи крові.
Частина еритроцитів
ІІІ. Осмотична резистентність. В середині еритроцитів вміст білків набагато вищий, ніж у плазми. Осмотиск в середині еритроцита трохи нижчий, ніж у плазмі. Це забезпечує нормальний тургор еритроцитів. Оскільки мембрана його для білків непроникна, основним компонентом, що забезпечує обмін води між еритроцитом і плазмою, є низькомолекулярні іони. Гемоліз відбувається в гіпотонічному розчині. Мірою осмотичної стійкості (резистентності) еритроцитів е концентрація NaCl в розчині, при якій відбувається гемоліз. У нормі гемоліз починається при 0,4% концентрації NaCl (мінімальна резистентність). При такій концентрації соляної кислоти руйнуються найменш стійкі еритроцити. У 0,34 % NaCl руйнуються всі еритроцити. При деяких захворюваннях осмотична стійкість еритроцитів зменшується, і гемоліз відбувається при вищій концентрації розчину NaCl. У гіпертонічному розчині із еритроцитів виходить вода, вони на певний час зморщуються.
ІУ. Гемоглобін (Нв). Головна функція еритроцитів – транспорт О2 і СО2. Транспорт кисню виконується в організмі практично тільки еритроцитами завдяки вмісту в них гемоглобіну. В крові його 14-16 %, а в окремому еритроциті 32%. Всього в організмі людини близько 750-800 г гемоглобіну. Якщо Нв буде в плазмі крові (гемоглобінемія), а не в еритроцитах, то він швидко виводиться через нирки, й токсично на них впливає (гемоглобінурія). Перебуваючи в плазмі, гемоглобін значно підвищить онкотичний тиск і в’язкість крові, а це ускладнює кровообіг.
Молекула Нв складається з двох частин – глобіну і небілкової частини – гема. Глобін становить 96% маси молекули. У молекулі Нв одна молекула глобіну і чотири молекули гема. До складу гема входить атом заліза, здатний приєднувати і віддавати молекулу кисню. При цьому валентність заліза не змінюється., тобто воно залишається двохвалентним.
Сполуки гемоглобіну:
Оксигемоглобін – НвО2. Сполука нестійка. Яскраво червоний колір.
Відновлений ННв (дезоксигемоглобін) – коли Нв віддав кисень тканинам.
Карбгемоглобін (НвСО2) – гемоглобін з вуглекислим газом. Таким чином глобін переносить від тканин до легень СО2.
Метгемоглобін (МtНв) – це сполука глобіну з киснем, в якому залізо стає тривалентне (під дією сильних окисників, отруєння аміл-пропілнітрітами).
Карбоксигемоглобін (НвСО)- це сполука з чадним газом. Досить стійка, віддає газ повільно, лише за умови вдихання чистого повітря.
В синтезі Нв грають роль вітаміни В6,12, фолієва кислота, залізо.
В нормі Нв здорового чоловіка міститься 130-160 г/л, у жінок – 120-140 г/л.
В скелетних м’язах знаходиться міоглобін, він зв’язує 14% кисню в організмі. Цей білок (з меншою молекулярною масою) постачає киснем тканини в умовах гіпоксії при напруженій роботі м’язів, а також відбирає О2 від Нв і передає його м’язовій тканині.
Колірний показник (КП) – відносний вміст Нв в одному еритроциті (в нормі 0,85-1,05).
У. Швидкість осідання еритроцитів (ШОЄ) – це здатність еритроцитів осідати.
У нормальних умовах еритроцити знаходяться в плазмі у вигляді емульсії. Якщо із судин взяти кров, додати до неї лимоннокислого натрію і помістити її в циліндр, то через деякий час можливо спостерігати, як еритроцити осідають на дно циліндра (вгорі прозорий шар). Під мікроскопом ці еритроцити мають вигляд так званих «монетних стовпчиків». ШОЄ залежить від білкового складу плазми: гамма-глобуліни і фібриноген збільшують, а альбуміни – зменшують ШОЄ. В нормі у чоловіків ШОЄ становить 1-10 мм/г, у жінок – 2-15 мм/годину. ШОЄ зростає при вагітності, мензіс (підвищуеться фібриноген), при запаленнях, онкологічних захворюваннях. ШОЄ визначають за допомогою апарата Панченкові.
Лекція №4.
Тема: Механізми збудження. Проведення збудження.
План:
І.Потенціал дії.
ІІ. Зміни збудливості клітини під час дії ПД.
ІІІ. Зміни МП при дії електричного струму.
ІY. Механізм проведення нервового імпульсу по нервовим волокнам.
І. Потенціал дії (ПД) або електричний імпульс –швидка високоамплітудна зміна потенціалу мембрани (МП) збудливої клітини при її збудженні.
Методи реєстрації потенціалу дії:
Позаклітинний (метод пошкодження при розриви органа) – досліджують нерв чи м’яз. Електроди прикладають до пошкодженої і непошкодженої поверхні, виникає різниця потенціалів.
А) біполярне дослідження реєструється двохфазний сумарний ПД (обидва електроди є активними, дві фази ПД відповідають руху хвилі збудження спочатку під одним, а потім під іншим електродом)
Б) уніполярна реєстрація - один електрод є активним, а інший – пасивним і реєструється однофазний сумарний ПД.
2. Внутрішньоклітинний (мікроелектродний) – реєструється ПД окремої клітини.
Фази ПД:
А) Фаза деполяризації ПД – МП різко зменшується до нуля. Виникає різниця потенціалів протилежного знаку.
Б) Фаза реверсполяризації (перезарядка мембрани) - заряд мембрани змінюється на протилежний –зовні «-», всередині «+». Реверсія МП – це кількість мВ, на яку ПД перебільшує МП. Натрієва активація.
В) Фаза реполяризації ПД (відновлення поляризації) –МП поступово повертається до вихідного рівня. Ця фаза довша за деполяризацію. Після закінчення піку ПД мембрана протягом15-30 мс залишається ще частково деполяризованою – слідовий негативний потенціал. Овершут – пік ПД. В основі – натрієва інактивація (припинення входу натрію в клітину) і калієва активація.
Г) Слідова гіперполяризація (=позитивна частина слідового потенціалу). Виникає внаслідок збільшення різниці між внутрішньою і зовнішньою мембранами (наприклад -95 мВ проти -90 мв).
В основі формування ПД лежать зміни проникності мембрани, що пов’язані із змінами стану каналів мембрани під впливом подразника. Подразник зумовлює відкриття натрієвих каналів і вхід натрію в клітину (іони натрію заряджені позитивно). Натрій нейтралізує негативний внутрішній заряд, і рівень поляризації мембрани зменшується – деполяризація. Незначний процес деполяризації, який виникає на початку збудження і не доходить до критичного рівня, називають місцевим потенціалом мембрани, або локальним потенціалом (ЛП). ЛП залежить від сили подразника; зникає після того, як припиняється незначне подразнення; зростає відносно повільно; здатний до сумації; нездатний до незатухаючого розповсюдження по волокну. Як тільки ЛП досягає певного критичного рівня (для нервового волокна 10мВ), починається деполяризація мембрани, що збільшує проникність іонів натрію, а це у свою чергу призводить до подальшої деполяризації, виникає вибухоподібний процес збудження.
Іонний механізм ПД.
При досягненні критичного рівня деполяризації настає швидке відкриття значної кількості натрієвих каналів – лавиноподібне проникнення натрію в клітину і повна деполяризація мембрани, яка супроводжується реверсією МП. Зовнішня частина мембрани заряджується негативно, а внутрішня – позитивно (інверсія фаз). При досягненні піка ПД рух натрію раптово припиняється, але продовжується вихід калія. Закінчується пік ПД, починається відновлення поляризації мембрани – реполяризація. В процесі реполяризації беруть участь калієві та натрієві насоси (натрій викачується назовні, потім калій вкачується всередину клітини). Калію надходить всередину клітини більше, ніж вийшло під час збудження - це гіперполяризація мембрани.
Параметри ПД:
1. Амплітуда – 100-120 мВ.
2.Протяжність – 1-3 мс в нервових волокнах, в м’язових довше ( в типових кардіоміоцитах – 300мс).
Фізіологічна роль ПД – полягає в тому, що він забезпечує збудження клітини та передачу цього збудження на інші клітини, а також підтримку чи активацію у збудливих клітинах специфічних функцій.
ІІ. При збудженні змінюється збудливість тканин.
Фази змін збудливості:
1)Початкове підвищення збудливості - під час розвитку локального потенціалу, пов’язане з підвищенням проникності мембрани для всіх іонів.
2)Рефрактерний період (зниження збудливості тканин) – під час піку ПД. Під час висхідної частини піку ПД збудливість зменшується до нуля (абсолютна незбудливість або абсолютна рефрактерність) – навіть над порогові подразники не збуджують тканину ( тому що вже відкрити всі натрієві канали). Відносна рефрактерність – поступове відновлення збудливості, але збудливість нижча за вихідний рівень (низхідна частина піку ПД), на надпорогові подразники можлива відповідь.
3) Супернормальний період (фаза екзальтації) - підвищення збудливості тканини у порівнянні з вихідною. В основі – слідові деполяризація мембрани (негативний слідовий потенціал). Тканина відповідає на подразник підпорогової сили.
4) Субнормальний період – збудливість незначно знижена (фаза співпадає з позитивним слідовим потенціалом, або з гіперполяризацією мембрани). Після закінчення цієї фази встановлюється початкова збудливість тканини.
Тривалість фаз збудливості для волокон рухового нерву:
абсолютна рефрактерність – 0,5 мс, відносна рефрактерність – 4,8 мс, супернормальна фаза – 15-30 мс, субнормальна – 50 мс.
Обов’язковими компонентами процесу збудження є:
Розвиток ПД на мембрані.
Виникнення специфічної функції (скорочення м’яза та ін.)
Показники збудливості:
Поріг сили подразника – мінімальна сила подразника, при якій він викликає збудження. Чим вища збудливість, тим менший поріг сили подразника. При необхідності оцінити рівень збудливості тканини в клінічній практиці визначають поріг сили подразника для цієї тканини.
Пороговий потенціал – різниця між МП та критичним рівнем деполяризації мембрани (Екр). Критичний рівень деполяризації – це той рівень МП, при зменшенні якого до МП на мембрані виникає ПД. Отже, пороговий потенціал – різниця між величиною МП та критичним рівнем деполяризації мембрани, показує на скільки мВ необхідно зменшити МП, щоб на мембрані виник ПД. Тобто, чим менший поріг деполяризації, ти м вища збудливість клітини та навпаки.
ІІІ. При дії на клітину електричного струму як подразника, збудження (ПД) виникає під катодом (катодний струм), при вхідному напрямку струму, так як саме під катодом виникає деполяризація мембрани. І, якщо ця деполяризація досягає критичного рівня, на мембрані виникає збудження, ПД. Збудження під катодом виникає, якщо сила струму , час дії збудника не нижчі порогових величин.
Закон полярної дії постійного струму (закон Пфлюгера) – при замиканні струму збудження виникає під катодом, а при розмиканні – під анодом. Під катодом позитивний потенціал на зовнішній мембрані зменшується – виникає деполяризація і потім збудження. Під анодом позитивний потенціал на зовнішньому боці мембрани збільшується – гиперполяризація мембрани і збудження не виникає. При найменший силі струму – скорочення тільки при замиканні струму на катоді, при збільшенні реобази (сили) – при замиканні і на аноді. Катодне замикательне скорочення більше анодного замикательного і розмикательного і катодного розмикательного скорочення.
ІY. Збудження виникає в певній частині мембрани і розповсюджується вздовж нервового або м’язового волокна. Це відбувається тому, що збуджена ділянка стає джерелом утворення місцевих колових струмів, тобто направленого потоку електронів між збудженою і ще не збудженою ділянками мембрани клітини. На поверхні волокна струм тече від не збудженої до збудженої ділянки, а всередині волокна – навпаки. Утворюється коло струму, яке пронизує мембрану на певній відстані від збудженої ділянки у висхідному напрямку. Такий трансмембранний потік викликає конформацію білків мембрани, що утворюють натрієві канали і трансмембранний потік натрію всередину мембрани в сусідній ділянці. За цим виникають деполяризація мембрани клітини і розвиток локального потенціалу. Коли деполяризація досягає критичного рівня, з’являється новий генеративний потенціал дії і процес збудження переміщується на наступну ділянку. Далі виникає ефекторний механізм: попереду хвилі збудження розповсюджується хвиля локального, або кільцевого струму, що перетинає незбудливу частину мембрани, розповсюджується на значну відстань від місця збудження і у віддалених частинах виникає посилення збудження, а у частині, що розташована поруч із збудженою ділянкою мембрани, розвивається нове збудження. Так ПД розповсюджується по всій поверхні нервового волокна, якої б довжини воно не було, без зниження своєї величини (120 мВ), тобто не затухаючи (бездекрементно).
У м’язових і не мієлінових волокнах збудження розповсюджується безперервно «від крапки до крапки», по мієліновому нервовому волокну – стрибкоподібно від одного вузла до іншого, а інколи перестрибують через 2-4 вузли. Такий тип проведення збудження називається сальтаторним. Воно можливе тільки тому, що кільцевий струм, який перетинає ще не збуджену мембрану, в 5 разів вищий, ніж порогів для виникнення ПД.Ця властивість кільцевого струму називається фактором гарантії. Швидкість сальтаторного розповсюдження значно вища, ніж безперервного, У сальтаторному воно досягає 120 м/с, а у безперервному нервовому волокні – 0,5 м/с.
Механізм проведення збудження по нервовим волокнам.
Під впливом П на мембрані волокна виникає збудження – ПД (зовні «-«, всередині «+», фаза реверсполяризації). Між збудженими та не збудженими (зовні «+», всередині «-» : потенціал спокою) ділянками мембрани волокна виникає різниця потенціалів. Зовні та всередині ці ділянки сполучені провідним середовищем (цитоплазма, міжклітинна речовина), і між цими ділянками виникають місцеві електричні струми, що спрямовані від «+» до «-», діють на мембрану із зовнішньої та внутрішньої поверхні. Ці струми є подразниками для мембрани. Струми мають вихідний (катодний) напрям в не збуджених ділянках мембрани. Отже, на збуджену ділянку мембрани нервового волокна діє катодний електричний струм, сила, час дії та швидкість збільшення сили, якого вищі порогу – цей струм викличе деполяризацію мембрани, ПД на мембрані не збудженої ділянки. Далі процес повторюється і ПД поширюється вздовж всієї мембрани нервового волокна.
Закони проведення збудження по нервовим та м’язовим волокнам.
Закон фізіологічної неперервності чи фізіологічної цілісності волокна – для здійснення проведення необхідним є нормальний функціональний стан мембрани волокна. Якщо його пошкодити, обробивши, наприклад, новокаїном, проведення припиняється (новокаїн інактивує натрієві канали мембрани).
Закон двостороннього проведення – в умовах експерименту збудження поширюється вздовж волокна в два боки, тобто провідність волокон двобічна. Але в умовах цілісного організму відбувається однобічне проведення збудження по нервовим волокнам, незважаючи на двобічну провідність – ПД поширюється від рецепторів до нервових центрів, від нервових центрів до органів-ефекторів. Це відбувається завдяки тому, що обов’язковим елементом будь-якої рефлекторної дуги є синапси, які мають однобічну провідність.
Закон ізолюванного проведення – ПД поширюючись по мембрані одного волокна, не передається на сусідні. Причина цього полягає в тому, що мембрана волокна має набагато більший опір, ніж міжклітинна рідина і місцеві струми шунтуються через міжклітинну рідину й не викликають ПД на мембрані сусідніх волокон.
Закон бездекрементного проведення – амплітуда ПД не змінюється при проведенні збудження по мембрані волокна. Причиною цього є те, що ПД підкоряється закону «все або нічого» (підпороговий подразник не викликає відповідної реакції, на пороговий подразник виникає максимальна реакція).
Проведення без розвитку втоми.
Проведення з дуже високим коефіцієнтом надійності.
Проведення з великою швидкістю.
ЛЕКЦІЯ № 4. Фізіологія м'язів.
1. Фізичні та фізіологічні властивості скелетних, серцевої і гладких мишц
За морфологічними ознаками виділяють три групи м'язів:
1) поперечно-смугасті м'язи (скелетні м'язи);
2) гладкі м'язи;
3) серцевий м'яз (або міокард) .
Опції поперечно-смугастих м'язів: 1) рухова (динамічна та статична);
2) забезпечення дихання;
3) мімічна;
4) рецепторна;
5) депонує;
6) терморегуляторная.
Опції гладких м'язів:
1) підтримка тиску в порожнистих органах;
2) регуляція тиску в кровоносних судинах;
3) спорожнення порожнистих органів і просування їх содержімого. Функція серцевого м'яза - насосна, забезпечення руху крові по сосудам.
Фізіологічні властивості скелетних м'язів:
1) збудливість (нижче, ніж у нервовому волокні, що пояснюється низькою величиною мембранного потенціалу);
2) низька провідність, порядку 10-13 м / с;
3) рефрактерність (займає за часом більший відрізок, ніж у нервового волокна);
скорочення. Одиночні скорочення виникають при дії одиночного роздратування, а титанічні виникають у відповідь на серію нервових імпульсів;
4) лабільність;
5) скоротність (здатність зменшуватися або розвивати напруга) .
Розрізняють два види скорочення:
а) изотоническое скорочення (змінюється довжина, тонус не змінюється);
б) ізометричне скорочення (змінюється тонус без зміни довжини волокна). Розрізняють одиночні і титанічні
6) еластичність (здатність розвивати напругу при розтягуванні) .
Фізіологічні особливості гладких мишц.
Гладкі м'язи мають ті ж фізіологічні властивості, що й скелетні м'язи, але мають і свої особливості:
1) нестабільний мембранний потенціал, який підтримує м'язи в стані постійного часткового скорочення - тонусу;
2) мимовільно автоматичну активність;
3) скорочення у відповідь на розтягнення; 4) пластичність (зменшення розтягування при збільшенні розтягування);
5) високу чутливість до хімічних веществам.
Фізіологічною особливістю серцевого м'яза є її автоматизм. Збудження виникає періодично під впливом процесів, що протікають в самій м'язі. Здатністю до автоматизму мають певні атипові м'язові ділянки міокарда, бідні і багаті миофибриллами саркоплазмою.
2. Механізми м'язового сокращенія
Електрохімічний етап м'язового сокращенія.
1. Генерація потенціалу дії. Передача збудження на м'язове волокно відбувається за допомогою ацетилхоліну. Взаємодія ацетилхоліну (АХ) з холінорецепторами призводить до їх активації і появі потенціалу дії, що є першим етапом м'язового сокращенія.
2. Поширення потенціалу дії. Потенціал дії поширюється всередину м'язового волокна по поперечної системі трубочок, яка є зв'язуючою ланкою між поверхневою мембраною і скорочувальним апаратом м'язового волокна.
3. Електрична стимуляція місця контакту призводить до активації ферменту і утворення інозілтріфосфата, який активує кальцієві канали мембран, що призводить до виходу іонів Ca та підвищення їх внутрішньоклітинної концентраціі.
Хемомеханіческій етап м'язового сокращенія.
Теорія хемомеханіческого етапу м'язового скорочення була розроблена О. Хакслі в 1954 р. і доповнена в 1963 р. М. Девісом. Основні положення цієї теорії:
1) іони Ca запускають механізм м'язового скорочення; 2) за рахунок іонів Ca відбувається ковзання тонких Актинові ниток по відношенню до міозіновим.
У спокої, коли іонів Ca мало, ковзання не відбувається, тому що цьому перешкоджають молекули тропоніну і негативно заряди АТФ, АТФ-ази та АДФ. Підвищена концентрація іонів Ca відбувається за рахунок надходження його з межфібріллярного простору. При цьому відбувається ряд реакцій за участю іонів Ca:
1) Ca2 + реагує з тріпоніном;
2) Ca2 + активує АТФ-азу;
3) Ca2 + знімає заряди з АДФ, АТФ, АТФ-ази. Взаємодія іонів Ca з тропонином призводить до зміни розташування останнього на актиновой нитки, відкриваються активні центри тонкої протофібрілли. За рахунок них формуються поперечні містки між актином і міозином, які переміщують Актинові нитку в проміжки між миозиновой ниткою. При переміщенні актиновой нитки щодо миозиновой відбувається скорочення м'язової ткані.
Отже, головну роль в механізмі м'язового скорочення грають білок тропонін, який закриває активні центри тонкої протофібрілли і іони Ca.
Лекція №5.
Тема: Фізіологія серця. Серцевий цикл.
План:
І. Серця. Загальна будова. Серцевий м’яз.
ІІ. Фізіологічні властивості міокарда.
ІІІ. Автоматизм серця. Провідна система.
ІУ. Серцевий цикл. Його фази.
У.Систолічний і хвилинний об’єм крові.
УІ. Електричні явища в серці. Серцеві тони.
І. Серця. Загальна будова. Серцевий м’яз. Рух крові забезпечується скороченням серця, яке нагнітає її в судинну систему. Серце складається з чотирьох камер: двох передсердь і двох шлуночків. Ліва і права частини серця відділені суцільною перетинкою, тому їх розглядають як різні відділи, що працюють одночасно. «Ліве серце» складається з передсердя і шлуночка і нагнітає кров у велике коло кровообігу. Праве передсердя і шлуночок («праве серце») забезпечують насосну функцію для циркуляції крові по малому колу кровообігу (легенева циркуляція). Передсердя мають тонку м’язову стінку, оскільки вони виконують функцію тимчасового резервуара крові, яка надходить, і проштовхують її до шлуночків. Міокард передсердь має два шари: циркулярний і повздовжній. Циркулярні волокна оточують переважно судини, що впадають в передсердя, а при скороченні перетискують їх отвори. Шлуночки мають товсту м’язову стінку, м’язи якої потужно скорочуються, що забезпечує проштовхування крові на значну відстань. Міокард шлуночків складається з трьох шарів: зовнішній і внутрішній шар - спіралеподібної форми, середній шар – з циркулярних волокон. Між передсердями і шлуночками є клапани, які сприяють руху крові тільки в одному напрямку. Вушки – додаткові ємкості на передсердях.
Будова міокарда. До складу міокарда входять кілька типів клітин, схожих за будовою, але різних за функціями: посмуговані м’язові клітини міокарда передсердь і шлуночків, які виконують функцію скорочення, та клітини провідної серцевої системи. М’язові клітини міокарда мають посмугованість внаслідок чергування світлих (ізотропних) і темних (анізотропних) ділянок, але на відміну від скелетних м’язових волокон серцеві міоцити мають бокові виступи, де розміщуються вставні диски( нексуси- щілиноподібні контакти), за допомогою яких міоцити з’єднуються один з одним і утворюють сітку (функціональний синцитій міокарда – коли збудження виникло в будь-якої ділянки, то воно розповсюджується на інші ділянки). Електрична взаємодія в кардіоміоцитах: ділянки нексусів мають низький електричний опір по відношенню з іншими частинами мембран, що забезпечує швидкій перехід збудження з одного волокна на інше. Скоротливими елементами серцевих міоцитів є актинові і міозинові міофіламенти. До складу міокардіоцитів входять
також саркоплазматична сітка (депо іонів кальцію), якої менше, чим у скелетних м’язів та ще інші органоїди.
ІІ. Фізіологічні властивості міокарда.
Серцевий м’яз за своїми функціональними властивостями займає проміжне місце між посмугованими і гладкими м’язами. Міокард подібний до посмугованих скелетних м’язів здатністю швидко та інтенсивно скорочуватися, а до гладких м’язів – здатністю до мимовільної активності і чутливістю до впливу вегетативних нервів і гормонів.
Міокард має такі фізіологічні властивості: збудливість, провідність, скоротливість, рефрактерність і автоматизм.
А)Збудливість. Серцевий м’яз менш збудливий, ніж скелетний. Тобто, у серцевого м’язу більш високий поріг подразливості.
Деполяризація мембрани починається при досягненні критичного рівня, який дорівнює -50- -55мВ. В скоротливих або типових кардіоміоцитах (Т-КМЦ) можливо виділити 5 фаз розвитку ТМПД( трансмембранного потенціалу дії): 0 – деполяризація, 1,2,3 – реполяризація, 4 – діастолічна деполяризація.
0 – швидка деполяризація (10мс) – обумовлена короткочасним підвищенням мембран для Nа + (спочатку канали відкриваються, а потім швидко закриваються): швидкий натрієвий струм. Також повільно в клітину входить кальцій. Досліди з використанням мікроелектородної техники показали, що при досягненні різниці потенціалів критичного рівня виникає різкий спад електричного заряду клітин. Це свідчить про їхнє збудження. Чим швидше настає критичний рівень деполяризації, тим частіше скорочується серце. Виникає перезарядка мембрани (реверсія): внутрішня поверхня мембрани має «+», а зовнішня – «-».
1 - фаза початкової (швидкої) реполяризації – інактивація натрієвих каналів обумовлена швидким струмом іонів хлору в клітину. Активація виходу іонів К із клітини. ТМПД =0.
2 - плато (повільна реполяризація) (200 мс) – повільний вхід іонів Са2+ , Nа+ в клітину (кальцієво-натрієвий механізм) і вихід із клітин іонів К. ТМПД підтримується на одному рівні (плато). Кардіоміоцити знаходяться в збудженому стані.
3 – фаза кінцевої (швидкої) реполяризації – інактивація вільних вхідних струмів Са2+, Nа+ при посиленому виходу К+. ПД повертається до спокою: до ТМПП. Відновлюється поляризація мембрани: зовнішня поверхня мембрани –«+», а внутрішня – «-».
4 –фаза діастоли – спонтанна діастолічна деполяризація формується повільним струмом: вихід К+; вхід Nа+, Са 2+ (працює Nа -К насос). ТМПД стає - -90мВ.
Клітини провідної системи серця володіють здатністю до спонтанного повільного збудження - зниження негативного заряду внутрішньої поверхні мембрани під час фази 4 – спонтанної діастолічної деполяризації, вона лежить в основі автоматичної активності клітин синусового вузла та провідної системи серця, тобто здатності до мимовільного зародження в них електричних імпульсів. Безперервна ритмічна активність серця протягом усього життя можлива лише у випадку ефективної роботи іонних насосів міоцитів вузлів.
Під час діастоли з клітини виводяться іони натрію, а в середину повертаються іони калію. Іони кальцію, які проникли в саркоплазму, повертаються в саркоплазматичну сітку. Ці процеси відбуваються з великою затратою енергії АТФ, КФ.
Міокард на порогове і надпорогове подразнення відповідає максимальним скороченням, сила його скорочення не залежить від сили подразнення ( а в скелетному м’язі: чим більша сила подразнення, тим більша сила скорочення ).
ПД серцевого м’язу суттєво відрізняється від скелетного:
коротка фаза деполяризації;
тривала фаза реполяризації.
Закон Боудича «все або нічого»: на подразнення (порогове, надпорогове) міокард відповідає збудженням усіх волокон (максимальне скорочення) або не реагує зовсім ( якщо подразник підпороговий). Отже, серце відповідає тільки на порогове подразнення, і відповідає по максимуму. Це відбувається завдяки вставним дискам (нексусам) - серцевий м’яз поводиться як функціональний синцитій: збудження, що виникло в якому-небудь відділі, поширюється на інші клітини (в нервовій і посмугованій м’язовій тканині клітини збуджуються ізольовано).
Б) Провідність серця. Хвилі збудження проводяться по різнім структурним елементам серця (провідної системі, скоротливому міокарду) з неоднаковою швидкістю. В кардіоміоцитах шлуночків швидкість проведення збудження становить 0,3 -1,0 м/с. Збудження, яке виникає в синусовому вузлі, проводиться по передсердям зі швидкістю 0,8-1,0 м/с. Далі імпульси розповсюджуються униз і вправо. Передача збудження з передсердь на шлуночки (верхня частина АВ вузла) виникає дуже повільно (0,002 м/с) – атріовентрикулярна затримка, завдяки якої систола передсердь відділяється від систоли шлуночків. По волокнам пучка Гіса, волокнам Пуркін’є збудження розповсюджується зі швидкістю 2-5 м/с (тому що тут є швидкі натрієві канали).
Причини великої швидкості проведення збудження по провідній системі серця є:
великий діаметр волокон;
наявність нексусів;
достатня амплітуда ПД (близько 100 мВ).
В) Скоротливість міокарда. Після виникнення ПД в м’язових волокнах передсердь і шлуночків вони скорочуються поодинокими, але тривалими скороченнями, що називають сполученням збудження і скорочення. При чому скорочуються не окремі волокна, а увесь міокард. Скорочення саркомера відбувається внаслідок втягування ниток актину між нитками міозину. Скорочення кардіоміоцитів починається, коли в саркоплазмі підвищується концентрація вільного кальцію (за рахунок вихіду із СПР та входу його із міжклітинної речовини крізь відкриті кальцієві канали). Під час ПД кальцій із системи Т-трубочок, де він знаходиться як клітинний резерв, входить в саркоплазму. Якщо під час досліду з розчину Рінгера усунути іони кальцію, то порушується взаємозв’язок між збудженням і скороченням. У відповідь на подразнення м’язового волокна скорочення не відбувається. Процес розслаблення міокарда починається внаслідок зв’язування іонів кальцію в саркоплазматичній сітці, а також перенесення іонів кальцію через сарколему в міжклітинну рідину.
Першими скорочуються м’язи передсердь, потім папілярні м’язи і субендокардіальний шар м’язів шлуночків. В подальшому скорочення охоплює і внутрішній шар шлуночків, забезпечуючи тим самим рух крові із порожнин шлуночків в аорту і легеневий стовбур. Серце для власної механічної роботи (скорочення) получає енергію, яка звільняється при розпаді АТФ. КФ.
Г) Рефрактерний період. В серці на відмінність від інших збудливих тканин є значно виражений і подовжений рефрактерний період (період зміни збудливості). Він характеризується різким зниженням збудливості тканин у перебігу її активності. Збудливість міокарда змінюється протягом всього періоду скорочення (тобто пов’язана з діяльністю ПД).
Розрізняють 3 фази збудливості:
Незбудлива фаза (абсолютної рефрактерності) – стан збудженого міокарда, коли він не відповідає на подразник будь-якої сили. Тривалість – 0,25 с, тобто триває майже весь період скорочення серця (систола і початок діастоли передсердь і шлуночків). Це пов’язано з інактивацією натрієвих каналів, відкриття яких необхідно для виникнення нового ПД, а також тривалої деполяризації мембрани за рахунок вхідного струму іонів кальція.
Фаза відносної рефрактерності (під час діастоли передсердь і шлуночків) – збудливість поступово починає відновлюватись. Тривалість – 0,03 с. Міокард скорочується на дуже сильні подразнень (надпорогові). Можуть виникати екстрасистоли.
Фаза підвищеної (супернормальної) збудливості або екзольтація –
збудливість стає вище початкової. Міокард скорочується на підпорогові
подразнення.
Рефрактерність запобігає виникненню надто частих повторних збуджень міокарда, що могли б призвести до послаблення нагнітаючої функції серця. Під час рефрактерності шлуночки встигають розслабитися і починають наповнюватися кров’ю.
Якщо нанести додаткове подразнення під час закінчення рефрактерного періоду, то серце відповідає на нього додатковим позачерговим скороченням – екстрасистолою. Оскільки після ЕС настає фаза незбудливості, то черговий імпульс від синусно-передсердного вузла не може викликати скорочення шлуночків, тому виникає компенсаторна пауза більш тривала, ніж у нормі. Екстрасистоли можуть з’явитися не тільки при штучному подразненні міокарда, але і під впливом різних патологічних процесів, при емоційному збудженні.
ІІІ. Автоматизм серця. Провідна система.
Автоматизм – здатність серця ритмічно скорочуватися під впливом імпульсів, які виникають в ньому самому. Серце має типові міозити (посмуговані, скорочувальні) і атипові (виникає і проводиться збудження). Атипові кардіоміоцити – А-КМЦ (пейсмекери або «водії»-клітини, де виникають ритмічні імпульси) згруповані в вузли в певних ділянках міокарда. Вони менші за розміри, ніж типові клітини серця, мають велику кількість взаємно переплетених виростів. В їхній структурі більше саркоплазми і мало міофібрил, мітохондрій, рібосом, слабко виражена Т-система саркоплазматичного ретикулуму. В вузлах, крім міоцитів знаходиться значна кількість нервових клітин і волокон.
Елементи ПС:
Синусовий (синусно-передсердний, S-A вузол, вузол Кіс-Флака ) вузол – у місця впадіння верхньої і нижньої порожнистої вен у праве передсердя. Вузол еліпсоподібний, довжиною 10-15 мм, шириною – 4-5 мм. Це – водій ритму, частота розрядів цього вузла близько 70 за 1 хв. Він пригнічує автоматизм усіх ділянок провідної системи, що розташовані нижче.
Передсердно-шлуночковий вузол (А-V вузол, антриовентрікулярний)– в товщі міжпередсердної перегородки на межі передсердь і шлуночків. Розмір – 7,5х3,5х1 мм. Тут виникають імпульси з частотою 40-50 за 1хв.
Пучок Гіса – проходить по міжшлуночковій перегородці. Він генерує імпульси з частотою 20-30 в 1 хв. Його довжина 12-40 мм, ширина 1-4 мм. Він ділиться на дві ніжки, які переходять в волокна Пуркіньє, які безпосередньо контактують з клітинами скоротливого міокарда.
Кожному відділу провідної системи притаманний автоматизм. При цьому зберігається градієнт автоматизму: зменшення здатності до автоматизму ділянок провідної системи, які більш віддалені від синусо - передсердного вузла.
Автоматичне виникнення збудження міоцитів вузлів пов’язане із спонтанним зменшенням потенціалу сарколем ( довільною діастолічною деполяризацією).
ПД А-КМЦ у синусо-передсердному вузлі.
В А-КМЦ є спеціальні потенціал-чутливі канали. В ході реполяризації мембрани КМЦ , МП досягає 60 мВ. В мембрані КМЦ:
Відкриваються повільні кальцієві канали, іони Са за градієнтом концентрації повільно входять в клітину,
Відкриваються повільні натрієві канали і іони Na за градієнтом концентрації повільно входять в клітину,
Закриваються калієві канали, знижується вихід К з клітини за градієнтом концентрації.
Стан каналів мембрани КМЦ веде до повільного зменшення МП (деполяризація мембрани). Ця деполяризація виникає без дії зовнішнього подразника (автоматично), і коли вона досягає критичного рівня (-45 мВ), виникає пік ПД. Ця частина змін в часі МП клітини, що володіє автоматією, є специфічною для неї і носить назву фаза повільної діастолічної деполяризації, або спонтанної деполяризації.
ІУ. Серцевий цикл. При частоті скорочення серця 75 ударів за 1 хвилину тривалість СЦ становить 0,8 с.
Механічна робота серця полягає в послідовному скороченні і розслабленні м’язових волокон. При цьому змінюється об’єм порожнин і кров нагнітається в судини. Скорочення передсердь і шлуночків називають систолою, а розслаблення – діастолою. Систола і діастола передсердь і шлуночків у нормальних умовах узгоджені між собою і складають єдиний цикл роботи серця. Скорочення міокарда супроводжується змінами тиску крові у порожнинах серця, артеріальних судинах, виникненням пульсової хвилі, звукових явищ тощо.
Розрізняють три фази роботи серця: скорочення передсердь, скорочення шлуночків, загальна діастола.
І. Систола передсердь і діастола шлуночків (0,1 с) – при скороченні передсердь передсердно-шлуночкові клапани відкриваються, і кров поступає в шлуночки ( доводячи об’єм крові в шлуночках до 110-140 мл).
ІІ. Систола шлуночків і діастола передсердь (0,3 с). Скорочення шлуночків починається з їхньої деполяризації, про що свідчить поява –QR комплексу ЕКГ. Скорочення шлуночків викликає підвищення кров’яного тиску. Півмісяцеві клапани аорти і легеневих артерій відкриваються, шлуночки спорожнюються через ці артерії.
Періоди систоли шлуночків:
1) Період напруження шлуночків ( до відкриття півмісяцевих клапанів)-0,08 с:
А) фаза асинхронного скорочення (0,05 с) – оскільки хвиля збудження поширюється по міокарду не відразу, м’язові волокна скорочуються асинхронно (неодночасно)- першими скорочуються кардіоміоцити біля провідної системи. Волокна, які скоротились, розтягують ще не збуджені волокна, що змінює форму шлуночка. Тиск крові у шлуночках у цей час дорівнює нулю.
Б) фаза ізометричного скорочення (скорочення м’язів шлуночків без зміни їхньої довжини)– виникає, коли збудження охоплює увесь міокард шлуночків (0,03 с). Тиск крові в шлуночках різко починає підвищуватись. У цю фазу замикаються передсердно - шлуночкові клапани. Клапани аорти і легеневого стовбура продовжують бути закрити (тому що тиск в цих судинах ще вищий, ніж у шлуночках). Таким чином, порожнини шлуночків закриті (тому що клапані закриті), і кров не дає можливості зменшити їхній об’єм. Фаза триває доти, поки тиск у шлуночках не перевищить тиск у аорті і легеневому стовбурі. Після цього клапани аорти і легеневого стовбура відкриваються і кров із шлуночків проштовхується в аорту і легеневі артерії.
2) Період виштовхування крові із шлуночків(0,25 с) - починається з відкриття клапанів аорти і легеневої артерії:
А) фаза швидкого вигнання (0,12 с) – коли скорочення шлуночків досягає максимального значення (тиск крові 120 мм рт. ст.. у лівому шлуночку і аорті та 25 мм рт.ст. у правому шлуночку і легеневій артеріі),
Б) фаза повільного вигнання (0,13 с)
ІІІ. Діастола шлуночків (0,4 с) – коли вигнання крові зі шлуночків
завершується:
А) Протодіастоліческій період (0,04 с) – до закриття півмісяцевих клапанів.
Б) Період ізометричного розслаблення (0,08 с) –коли всі клапані закрити. Після спорожнення шлуночки розслаблюються, півмісяцеві клапани закриваються зворотнім током крові в судинах. Серце залишається в фазі спокою до тих пір, доки кров, яка заповнює передсердя, не натисне на атріовентрикулярні клапани.
В) Період наповнення шлуночків - відкриваються атріовентирикулярні клапани і кров надходить з передсердь у шлуночки. Повністю шлуночки наповнюються кров’ю під час систоли передсердь.
В працюючому серці діастола передсердь частково співпадає з діастолою шлуночків - це називається пауза. В період паузи передсердя наповнюються кров’ю. За паузою йде систола передсердь.
У. Систолічний та хвилинний об’єм серця.
Кількість крові, що викидається шлуночком серця при кожному скороченні, називається систолічним об’ємом серця (СОС), або ударним об’ємом (УО). В середньому він становить 60-70 мл крові. Кількість крові, що викидається правим і лівим шлуночками, однакова.
Знаючи частоту серцевих скорочень (ЧСС) і систолічний об’єм, можна
визначити хвилинний об’єм серця (ХОС, або кровообігу=ХОК), або серцевий
викид: ХОС =СОС х ЧСС.У стані спокою у дорослої людини ХОС в середньому
складає 5л. При фізичних навантаженнях СОС може збільшуватися вдвоє, а
серцевий викид досягати 20-30 л.
Серцевий індекс – показник функції серця (гемодинаміки). Це відношення хвилинного об’єму серця до площі поверхні тіла (л/ (хв. х м2)):
СІ =ХОС/S. S - це площина тіла, вона розраховується за спеціальними номограмами. Наприклад, у людині, масою тіла 65 кг і зростом 165 см, S=1,6.
УІ. Електричні явища в серці. Діяльність серця супроводжується електричними явищами. Всі збудливі тканини в спокої мають позитивний електричний заряд. При виникненні збудження заряд збудженої ділянки змінюється на від’ємний. Це відбувається і в серці: при збудженні виникає різниця потенціалів – струм дії. Збудження, що виникло в серці, поширюється. Таким чином, все нові й нові ділянки стають електронегативними, а отже, і в нових ділянках виникає різниця потенціалів. Серце стає могутнім електрогенератором. Тканини тіла, маючи здатність до електропровідності, дозволяють реєструвати електричні потенціали серця з поверхні тіла. Методика дослідження електричної активності серця отримала назву електрокардіографії, а крива, що відтворює виникнення, поширення і стихання процесу збудження в серці, називається електрокардіограма (ЕКГ). Для запису діяльності серця людини використовують електрокардіограф. При цьому використовують не менше 12 –ти відведень. Існує три стандартних відведення від кінцівок, при яких два активних електроди накладають на кінцівки.
ЕКГ у всіх здорових людей є постійною і характеризується наявністю 5
зубців, які позначаються буквами P, Q, R, S, T. Зубці Р, R, Т – позитивні, QS –
негативні.
Зубець Р відповідає збудженню передсердь (сума потенціалів правого і лівого передсердь). Амплітуда його менша або дорівнює 0,2 мВ, а тривалість – 0,11 с.
Інтервал Р- Q – передсердно-шлуночковий інтервал, поширення збудження по провідної системі: характеризує швидкість розповсюдження збудження від вузла-водія до шлуночків (час розповсюдження деполяризації передсердь, АВ вузла, атріовентрикулярної затримки). Його тривалість 0,1-0,21 с.
Комплекс Q R S – до зубця Т. Він відображає деполяризацію і реполяризацію шлуночків (весь період збудження шлуночків) і зветься електрична систола шлуночків. Тривалість – 0,35-0,40 с при ЧСС=75 в хв..
зубець Q – початкова деполяризація (збудження) провідної системи шлуночків (міжшлуночкової перегородки, верхівки серця). Тривалість 0,04 с. Він незначний, може навіть не визначатися на ЕКГ.
зубець R – самий високий зубець, головний вектор, відображає подальше збудження по міокарду двох шлуночків: від ендокарда до епікарда (крім основи лівого шлуночка). Тривалість 0,04 с.
зубець S – кінець збудження шлуночків(деполяризація правого шлуночка); виникає в результаті різниці потенціалів між незбудженою основою лівого шлуночка і збудженим міокардом.
Зубець Т – реполяризація міокарда шлуночків. Тривалість – 0,16-1,24 с. Він виникає оскільки збудження зникає із міокарда неодночасно, виникає знов різниця потенціалів, тому що деякі ділянки міокарда вже реполяризовалися, а деякі – ще ні. Висота зубця відображає стан обмінних процесів в міокарді.
Інтервал Т-Р – характеризує відсутність різниці потенціалів в серці (загальна пауза). Цей інтервал є ізоелектричною лінією.
Навіть незначні зміни в нормальній діяльності серця відображається змінами на ЕКГ. Тому цей метод широко використовують при діагностиці патологічних змін у серці.
Робота серця супроводжується характерними звуками, які називаються тонами серця. Розрізняють чотири тони серця. Два з них (перший і другий) можна прослухати за допомогою фонендоскопа. Фонендоскоп – це невелика капсула, обтягнута мембраною. Від капсули відходять гумові трубки з наконечниками. Усі чотири тони можна зареєструвати на фотопапері, який рухається, якщо до грудей обстежуваного прикласти мікрофон, з’єднаний з осцилографом. Ця методика називається фонокардіографія. І тон називається систолічним – виникає під час систоли шлуночків (робота атріовентрикулярних клапанів, вібрація стінок шлуночків). Він протяжливий, низький. Вислуховується на верхівці серця. ІІ тон – діастолічний (під час діастоли шлуночків). Формується при закриванні клапанів аорти і легеневого стовбура. ІІІ тон виникає внаслідок вібрації стінок шлуночків у фазу швидкого заповнення їх кров’ю. ІУ тон – при систолі передсердь і поверненні частини крові в передсердя. При вадах клапанів тони змінюються – змінюється інтенсивність звуків, з’являються шуми.
Лекція №6.
Тема: Регуляція діяльності серця. Методи дослідження діяльності серця.
План: І. Регуляція серцевої діяльності: міогенна, нервова, гуморальна.
Механізм впливів ВНС на фізіологічні властивості серцевого м’яза.
ІІ. Механізм гуморального впливу на діяльність серця .
І. Регуляція серцевої діяльності:
міогенна – пов’язані з фізіологічними властивостями власно структур серця. Забезпечує рівність притоку крові по венам і її викиду в артерії.
нервова – здійснюється за допомогою інтра- і екстракардіальної системи
гуморальна – вплив різних фізіологічно активних речовин, що виробляються безпосередньо в серці і організмі.
1) Міогенні (внутрішньосерцеві) механізми регуляції.
А) гетерометрична саморегуляція («різний», «вимірювання») –відбувається у відповідь на зміну вихідної довжини волокон міокарда (сила скорочення міокарда залежить від її вихідної довжини волокон - розтягнута клітина скорочується з більшою силою). Гетерометрична СР забезпечує пристосування роботи серця до кількості крові, яка притікає до нього.
Закон серця ( Франка – Стерлінга) – сила скорочення міокарда перебуває у прямо пропорційній залежності від вихідного розтягнення м’язових волокон під час діастол (сила скорочення серця тим більша, чим більше кінцево-діастолічний об’єм камер серця –але до 180 мл, при подальшому збільшенні об’єму зменшується сила скорочення: негативний інотропний =скорочувальний ефект). Встановлена ультраструктурна основа цього закону: кількість актиноміозинових мостиків максимально зростає при розтягненні сакркомера до 2,2 мкм. При розтягненні в кожній міофібрилі актинові нитки інтенсивніше висовуються між міозиновими голівками, що призводить до зростання кількості резервних містків, тобто тих точок, які з’єднують актинові і міозинові нитки під час скорочення. Більш сильне розтягнення міокарда під час діастоли відповідає посиленому притоку крові до серця. Отже, чим більше м’язове волокно міокарда розтягнеться під час діастоли, тим більшу силу воно може розвинути при скороченні під час систоли. Тому серце перекачує в артеріальну систему стільки крові, скільки її надходить з вен.
Б) Гомеометрична саморегуляція («подібний», «вимірювання») – коли не має значення ступень розтягнення волокон міокарда при діастолі (ізометричний режим – стала довжина волокон) і здійснюється завдяки особливостям міоцитів.
Ефект Анрепа - при збільшенні тиску в аорті автоматично збільшується сила серцевих скорочень.
Це пов’язане з двома механізмами:
- збільшення кінцево – систолічного об’єму (систолічний об’єм крові автоматично збільшується при підвищенні частоти серцевого скорочення – ХОС може бути збільшеним до 15 л/хв),
- поліпшення живлення міокарда через коронарні судини.
Це сприяє збільшенню сили скорочення міокарда під час м’язової роботи, коли збільшується ЧСС і підвищується АТ.
Внутрішньоклітинні механізми забезпечують також тривале пристосування міокарда до підвищених фізичних навантажень, які діють систематично. Воно полягає в прискоренні синтезу скоротливих білків міофіламентів і розвитку структур, які забезпечують енергетичні потреби. При підвищеному м’язовому навантаженні спостерігається фізіологічна гіпертрофія міокарда.
В) Хронотропія. Залежність сили скорочення серця від ЧСС (драбина Боудіча). Серце у відповідь на підвищення ЧСС реагує збільшенням сили скорочень і, навпаки, зі зменшенням ЧСС сила скорочень падає. Механізм цього феномена зв’язаний з накопиченням чи падінням в біоплазмі концентрації кальцію, а також збільшенням чи зменшенням кількості поперечних містків, що зумовлює позитивні або негативні ефекти серця.
2) Нервова регуляція.
А) Інтракардіальна НС . В серці є нервові інтрамуральні сплетення, які забезпечують місцеві кардіо-кардіальні рефлекси. В сплетеннях існують всі види нейронів – аферентні, проміжності, еферентні, гальмівні, що забезпечують замикання рефлекторної дуги на рівні інтрамуральних гангліїв серця. В передсердях і лівому шлуночці е рецептори, які реагують на активну напругу і пасивне розтягнення серцевого м’яза. Н-д, при збільшенні притоку крові до правого передсердя збільшується скорочення лівого шлуночка.
Місцеві єферентні нейрони бувають двох типів – холінергічні (менш збудливі)і адренергічні (більш збудливі). При малому кровонаповнення шлуночків в рецепторах розтягнення виникає мала частота імпульсації ПД, яка активує тільки адренергічні еферентні нейрони і не збуджує холінергічні. В результаті виникає рефлекторне збільшення сили і частоти серцевих скорочень. Це збільшує серцевий викид і попереджає зниження АТ. При занадто великому кровонаповненні шлуночків, частота ПД збільшується, збуджуються холінергічні нейрони, а адренергічні гальмуються (механізм зворотного гальмування). В результаті при великому кровонаповнення шлуночків проявляється негативний хроно- і ізотропний ефекти. Це захищає судини від занадто великого викиду крові під час систоли, від їх розриву.
Б)Екстракардіальна НС- забезпечується імпульсами із ЦНС по блукаючому і симпатичним нервам.
Ядра блукаючого нерва розташовані в довгастому мозку, аксони закінчуються в інтрамуральних гангліях серця, потім відростки йдуть до провідної системи, міокарду, коронарних судин. Правий блукаючий нерв іннервує праве передсердя, передні відділи шлуночків, СА-вузол; лівий – АВ-вузол, задні відділи шлуночків.
Закон серцевого ритму ( закон Пейнбріджа)– чим більше приток крові (особливо в правому передсерді), тим більша сила і частота серцевих скорочень. Механізм – збудження механорецепторів, така велика частота гальмує імпульси вагусного нерва, йде активізація симпатичних нервів.
Ядра СНС розташовані в бічних рогах п’яти верхніх сегментах грудного відділу СМ. Їх відростки закінчуються в шийних і верхніх грудних симпатичних вузлах; далі відростки йдуть до серця. СНС іннервує провідну систему, міокард. Симпатичні і парасимпатичні нерви в міокарді утворюють замість синапсів термінали. Термінали нервів тісно переплетені, їх медіатори можуть оказувати модулюючий ефект на виділення один одного.
Вплив СНС - позитивний хронотропний (збільшення ЧСС), батмотропний (збудливість), дромотропний (провідність), ізотропний ефект (сила скорочень) і трофічний вплив.
Вплив блукаючого нерва – негативний ефект за тими ж самими критеріями. Правий нерв – здебільше на ЧСС, а лівий – на атріовентрикулярне проведення. При сильному подразненні блукаючого нерва не тільки гальмується робота серця, але й може настати його зупинка в діастолі (гіперполяризація мембран міокарда). При тривалому подразненні нерва виникає ефект вислизання серця з-під впливу блукаючого нерва (скорочення відновлюється). Імпульси з нервових закінчень передаються на серце через медіатори (ПСНС ацетілхолін, СНС норадреналін).
Центри ВНС знаходяться в постійному тонусі, який підтримується притокам імпульсів від рецепторів тіла (тонус блукаючого нерва більший).
В) Рефлекторна регуляція діяльності серця. Подразнення майже всіх рецепторів тіла змінює діяльність серця. Однак, провідними в цьому є рефлексогенні зони – скупчення закінчень чутливих нейронів в самої серцево-судинної системі: хемо- і барорецептори. Від цих зон (дуга аорти, розгалуження сонних і легеневих артерій) починаються рефлекси, які регулюють усю гемодинаміку. Подразнення барорецепторів цих зон виникає при підвищенному АТ. Рефлекси дуги аорти замикаються через центри блукаючого нерва, тому знижують ЧСС і силу скорочень.
Існують рецептори позасудинні – в органах, які змінюють роботу серця. Так, при сильному ударі в живот можливо виникнення остановки серця через сильний потік імпульсів від кишок до блукаючого нерва, різке підвищення його тонуса.
Велике значення в регуляції роботи серця мають підкоркові структури – гіпоталамус (модулююча дія на ВНС), лімбічна система емоції впливають на серце), рухові центри ЦНС (рухи впливають на скорочення серця), а також кора (умовнорефлекторна регуляція роботи серця через пам’ять).
ІІ. Механізм гуморального впливу на діяльність серця.
А) Вплив інкреторної функції серця – у серці (передсерді) утворюються біологічно активні сполуки (дигіталісоподібні чинники, катехоламіни, продукти арахідонової кислоти) і гормони, зокрема передсердний натрійуретичний та ренін-ангіотензинові сполуки. Обидва гормони беруть участь у регуляції скоротливої активності міокарда,ХОС. При дії на рецептори ренін-ангіотензинові сполуки розвивається гіпертрофія міокарда.
Б) Вплив іонів на функції серця. Мембрани відповідають на проникнення іонів. Роль в трансмембранному обміні речовин належить концентраційному градієнту, який визначається концентрацією їх у крові, в міжклітинній рідині. Збільшення зовнішньо клітинної концентрації іонів призводить до зростання пасивного надходження їх у кардіоміоцити, зниження – до «вимивання».
вплив Са 2+. Якщо рівень Са у крові знижується, то збудливість і скоротливість серця зменшується, а при збільшенні, навпаки, підвищується. При надлишку кальцію серце може зупинитися в систолі. Механізм цього явища зв’язаний з із рівням кальцію у клітинах провідної системи та робочого міокарда.
вплив К+. При зменшенні концентрації К+ у крові зростають пейсмекерна активність та ЧСС. При збільшенні, навпаки. Дворазове підвищення вмісту К+ в крові може призвести до зупинки серця в діастолі. Механізм цих змін пов’язаний зі зменшенням співвідношення між зовнішнім і внутрішньоклітинним К+, підвищенням проникності мембран до К+, зниженням потенціалу спокою (цей ефект використовують при хірургічних операціях на серці).
вплив Na+. Зниження вмісту натрію в крові може призвести до зупинки серця. В основі цього впливу – порушення градієнтного транспорту натрію і кальцію й поєднання збудливості із скоротливістю. Незначне підвищення натрію призведе до збільшення скоротливості міокарда.
В) Вплив гормонів: справжніх (адреналін, норадленалін, глюкагон, інсулін) і тканинних (ангіотензин ІІ, гістамін, серотонін). Механізм дії норадреналіну, гістаміну, серотоніну пов’язаний із відповідними рецепторами: в-адренорецепторами, Н2-гістаміновими і серотоніновими. При цьому йде активізація кальцієвих каналів, накопичується внутрішньоклітинний кальцій, що зумовлює підвищення діяльності серця. Адреналін, тироксин і глюкагон діють на міокард опосередковано, через активізацію утворення АТФ, це причина позитивної ізотропної реакції (посилення роботи міокарда). Ацетилхолін і брадикінін інактивує кальцієві канали – негативний вплив на функції серця. Ще ацетилхолін збільшує проникність мембрани для калію і виникає гіперполяризація мембрани, в результаті цього зменшується швидкість деполяризації, скорочується тривалість ПД, знижується сила скорочення.
Г) Вплив метаболітів. Для нормальної роботи серця потрібна енергія. Тому всі зміни вінцевого кровоточу, трофічні функції крові позначаються на роботі міокарда.
При гіпоксії, внутрішньоклітинному ацидозі на мембрані клітин серця блокуються повільні Са2+ -канали, що пригнічує скоротливу активність. Не витрачена АТФ на скорочення серця, забезпечує життєдіяльність серця. Збільшення в серці концентрацій КФ, вільних жирних кислот, молочної кислоти супроводжується підвищенням діяльності серця.
Лекція №7.
Тема: Фізіологія кровоносних судин.
План:
І. Основні закони гемодинаміки. Загальний периферичний опір судин.
Лінійна та об’ємна швидкість руху крові. Час повного круговороту крові.
ІІ. Кров’яний тиск: артеріальний, капілярний, венозний.
Фізіологічні основи вимірювання кров’яного тиску.
ІІІ. Артеріальний пульс, його основні параметри.
ІУ. Функціональна класифікація кровоносних судин.
І. Основні закони гемодинаміки.
Наука, що вивчає рух крові по судинах зветься гемодинаміка. Вона базується на законах гідродинаміки, на основі різниці гідростатичного тиску - кров рухається із ділянки високого тиску в ділянку з низьким тиском. Отже, умова кровообігу – різниця тиску, яка проштовхує кров по судинах і повертає її до серця. За її законами рух крові визначається градієнтом тиску - різницею між тиском в лівому шлуночку при його систолі та тиском в правому передсерді при його діастолі. Н-д, в кінці систоли в лівому шлуночку створюється тиск величиною 120-130 мм рт.ст., в цей час в правому передсерді, що знаходиться в діастолі, тиск падає до 0. Таким чином виникає градієнт тиску величиною 120-130 мм рт.ст.
Рух крові по судинах залежить від загального периферичного опору судин.
Периферичний опір складається з опору кожної окремої судини. Таким чином, всю систему судин можна уявити як одну судину, опір кровотоку якої визначається за формулою Пуазейля:
R= 8 * l *h /p * r4;
8-коефіціент пропорційності; l- довжина трубки h–в’язкість крові; r - радіус судини.
Однак розрахунки гемодинаміки за цією формулою неможливо, оскільки в організмі внаслідок скорочення м’язів безперервно змінюється геометрія судин. Крім того, в’язкість крові в судинах різного діаметра неоднакова. Чим менший діаметр судин, тим менша в’язкість крові ( в дрібних судинах біля стінки знаходиться рідина, а не клітини – опір рідини менший від опору клітин).
Отже, опір судин знаходиться в прямо пропорційній залежності від в’язкості крові і в обернено пропорційній – від радіуса судини.
Основний опір рухові крові виникає в артеріолах. При збільшенні опору в артеріолах відтік крові з артерій зменшується і тиск в них підвищується. Зниження тонусу артеріол збільшує відтік крові з артерій, що призводить до зменшення артеріального тиску. Зміна їх просвіту є головним регулятором рівня загального артеріального тиску.
Лінійна швидкість кровотоку –це швидкість руху частинок крові уздовж судини. Цей гемодинамічний показник залежить від площі поперечного перерізу судини (сумарного діаметру судин): чим більша площа перерізу (діаметр), тим повільніше рухається кров. В аорті (площа перерізу 2,5 см2) в систолу лінійна швидкість найбільша – 50 см/с, в капілярах (сумарна поперечна площа 2500 см2)– дуже мала: 0,5 мм/с. Діаметр аорти в 500 разів менший загального діаметра капілярів.
Об’ємна швидкість кровотоку – це кількість крові, яка протікає через поперечний переріз судини (або групу судин) за одиницю часу. Об’єм крові, що протікає через кожний відділ судинного русла за одиницю часу, завжди однаковий, тобто за 1 хвилину через аорту або легеневі артерії, або сумарний поперечний розріз на будь-якому рівні артерій, вен, капілярів, протікає однакова кількість крові (хвилинний об’єм крові). Він забезпечує постачання кисню та поживних речовин, звільнює тканини від кінцевих і деяких проміжних продуктів обміну. У стані спокою скелетні м’язи одержують лише 15-20% крові з об’єму серцевого виштовху, тоді як печінка, нирки і головний мозок – більше половини. Кількісне порівняння кровопостачання окремих органів одержують при визначенні кровотоку на 100 г маси органа за 1 хв. Найменший кровоток у шкірі, м’язах, найбільший – в ендокринних залозах, нирках, слинних залозах, у міокарді.
Час кровообігу крові – це час, необхідний крові для її переміщення по великому та малому колам кровообігу. У спокою цей час складає 20-23 с (23 систоли). 1/5 цього часу припадає на велике коло кровообігу.
ІІ. Тиск крові – один з основних показників гемодинаміки. Тиск дорівнює відношенню сили, з якою кров діє на стінки судини, до його площини. Величина кров’яного тиску (Р) прямо пропорційно залежить від об’єму крові, яка проходить через цю судину (за 1 хв) і величини периферичного опору.
Р= Q * R
Q – об’ємна швидкість кровотоку
R – гідродинамічний опір руху крові
Величина Q залежить від діяльності серця (ЧСС и сили СС).
Н-д, при послабленні серцевої діяльності або при крововтраті величина кров’яного тиску знижується.
Величина судинного опору залежить від довжини судини, в’язкості крові і ширини просвіту судини. З цих трьох величин найбільш регульованою є просвіт судин – при розширенні судин кров’яний тиск знижується. Найбільш високим опором володіють дрібні артерії й артеріоли, тому від їх просвіту залежить величина кров’яного тиску в першу чергу.
Величина кров’яного тиску залежить не тільки від тиску крові всередині на стінку судини (внутрішній тиск), але і від зовнішнього тиску оточуючих тканин. Різниця між цьома величинами складає трансмуральний тиск. Внутрішній тиск складається з гідродинамічного (джерело тиску – скорочення серця) і гідростатичного тиску крові.
Гідростатичний тиск - є наслідком дії сил гравітації на кров, яка знаходиться в судині. Цей тиск змінюється при зміні положення тіла в просторі. При стоянні гідростатичний тиск впливає на трансмуральний тиск, трансмуральний тиск знижується на 30 мм рт.ст. Чим нижче від серця знаходиться судина, тим більшим стає трансмуральний тиск. На рівні 120 см нижче серця трансмуральний тиск вище гідродинамічного десь на 90 мм рт.ст.
Розтягуючий тиск крові крові створює в стінці судин напругу. Взаємозв’язок напруги (F) і трансмурального тиску (Pt) виражається законом Лапласа:
Рt = F : r, де r - радіус судин.
Отже, чим менший радіус судин, тим менша напруга в його стінці при однаковому трансмуральному тиску. Завдяки цьому самі маленькі судини – капіляри не розриваються, не зважаючи на достатньо високий тиск крові на їхню стінку.
Кров’яний тиск поділяється на артеріальний, венозний і капілярний. Артеріальний тиск поділяється на:
Систолічний тиск Рс –тиск в аорті під час систоли лівого шлуночка. На максимумі вигнання крові = 120-130 мм рт.ст. Залежить від серцевого викіду і еластичності аорти.
Діастолічний тиск Рд – складає в аорті 80 мм рт.ст. Визначається кількістю крові, яка залишилася в судині попереду надходженням у нього нової порції. Залежить надходження крові від опору нищерозташованих резистивних судин.
Пульсовий тиск (Рп) – різниця між систолічним і діастолічним тиском. В аорті =40 мм рт.ст. Пульсовий тиск в малих судинах зростає (зростає Рс, знижується Рд). На пульсовий тиск впливають – систолічний об’єм крові й ступень розтягнення стінки судини. При великому систолічному об’ємі АД значно коливається під час систоли і діастоли серця. Чим більше крові виштовхується з кожною систолою в артерії, тим більше зростає в них тиск під час систоли і більше знижується під час діастоли. Між еластичністю (розтягуванням) стінки і Рп є зворотній зв’язок. Чим більш еластичні судини, тим більше крові може поступити в них без значного підвищення АТ. Тому у людей з еластичними артеріями систолічний викид викликає відносно менше підвищення АТ. При розвитку склеротичних змін, під час систоли судина погано розтягується, значно підвищується систолічний тиск і також Рп.
Середній артеріальний тиск – тиск за час всього серцевого циклу. Для плечової артерії Рср = Рд +1/3 Рп =90 мм рт.ст.
Середній пульсовий тиск має велике значення, оскільки визначає об’ємну швидкість кровотоку в системній циркуляції.
Кров’яний тиск від артеріального до венозного русла судинної системи знижується нерівномірно, що залежить від загального діаметру судин. У дорослої людини в аорті середній тиск дорівнює в середньому 100 мм рт.ст., у середніх артеріях доходить до 85 мм рт. ст., в артеріолах до 30 мм рт.ст, в капілярах 10-15 мм рт.ст. У венах тиск знижується до нуля, а в порожнистих венах під час вдиху він навіть нижчий від атмосферного тиску.
Кров’яний тиск збільшується з віком (після 40 років), у людей з підвищеною масою тіла АТ вищий, ніж у людей з нормальною масою. При тривалому курінні систолічний тиск зростає на 10-20 мм рт.ст., а при емоційному стресі – ще більше. Коливання АТ залежить від генетичних факторів. Схильність до гіпертонії передається спадково.
Вплив фізичної роботи на АТ:
Велика фізична робота, у спортсменів систолічний тиск збільшується до 200-240 мм рт.ст. Діастолічний тиск знижується внаслідок розширення судин скелетних м’язів.
Середньої важкості робота та легка фізична робота – систолічнимй тиск підвищується, що забезпечує достатнє кровопостачання м’язів. Діастолічний тиск майже не змінюється.
Для вимірювання АТ використовують метод, запропонований російським лікарем Н.С.Коротковим. При цьому використовують тонометр. Він складається з порожнистої гумової манжети, гумової груши, манометра.
ІІІ. Артеріальний пульс – це ритмічне коливання стінок артерій у зв’язку з діяльністю серця.
Пульсова хвиля поширюється по артеріях з великою швидкістю – 9 м/с, потім поступово слабшає і закінчується в капілярній сітці. Артеріальний пульс свідчить про нагнітальну функцію серця. Визначаючи властивості пульсу, ми можемо скласти деяку уяву про роботу серця.
Методи визначення АП:
1)Пальпаторне дослідження – на променевій артерії, притиснувши її до променевої кістки. Можна визначити пульсацію і інших поверхневих артерій: скроневої, пахвинної, сонної.
2) Сфігмограма – графічний запис пульсу. На неї розрізняють анакроту (відповідає систолі лівого шлуночка), катакроту (при зниженні тиску в аорті під час діастоли лівого шлуночка), дикротичний підйом (при зниженні тиску крові в шлуночках під час їх діастоли, кров з аорти вдаряє в півмісяцеві клапани, які вже закрити).
Властивості АП (визначаються пальпаторно):
Частота пульсу – це кількість пульсових коливань стінки артерії за 1 хв. В нормі вона звичайно співпадає з ЧСС (60-80 ударів/хв.).
Ритм пульсу – визначають за тривалістю інтервалів між сусідніми пульсовими коливаннями. Якщо ці інтервали однакові, говорять про ритмічний пульс, якщо ні – аритмічний пульс.
Напруженість пульсу – визначають тією силою, яку необхідно прикласти до артерії, щоб зупинити пульсову хвилю. Напруженість пульсу відтворює такий гемодинамічний показник. як артеріальний тиск.
ІУ. Функціональна класифікація судин.
Загальна будова стінки:
1)Внутрішня оболонка - ендотелій, що побудований з одного шару клітин. 2) М’язова оболонка - гладенькі м’язи, еластичні і колагенові волокна. М’язова оболонка створює активне напруження стінки судини (судинний тонус) і змінює її просвіт.
3)Зовнішня оболонка – із сполучної тканини, має власні судини, нерви.
Функціональна класифікація судин.
Амортизуючи судини (аорта,легеневий стовбур, великі артерії) – завдяки великому діаметру вони створюють незначний опір крові, вони можуть значно розтягуватися і вміщувати значну порцію крові. Ці пружні судини забезпечують безперервний, рівномірний рух крові по судинах.
Резистивні судини (судини опору) - кінцеві артерії, артеріоли, капіляр, венули.Особливо великий опір крові в артеріолах, діаметром 0,06-о,1 мм. Завдяки м’язовим волокнам вони можуть вони активно змінюють свій просвіт і регулюють ступінь кровопостачання відповідної ділянки тіла, а також гідростатичний тиск у капілярах цієї ділянки.
Судини-сфінктери (пре капілярні сфінктери) – це скупчення гладеньких м’язових клітин на початку капілярі. Вони регулюють кількість відкритих капілярів, тобто площу обмінної поверхні.
Обмінні судини – капіляри, через стінки яких проходять дифузія і фільтрація (обмін речовин між кров’ю і тканинами).
Ємкісні судини – вени, які здатні значно розтягуватися, можуть вміщувати або викидати великий об’єм крові без істотних змін параметрів кровотоку. У венах знаходиться до 75 % ОЦК, тоді як артеріальна система і серце вміщують до 20%, а капіляри 5% ОЦК. Таким чином вени виконують функції резервуара перемінного об’єму крові
Шунтуючі судини – це артеріально-венозні анастомози, по яких кров переходить з артеріол у венули, минаючі капіляри.
Капіляри
Середній капіляр має радіус від 6 до 2—3 мкм, довжину — 750 мкм. При площі поперечного перерізу до ЗО мкм2 обмінна площа капіляра складає близько 14 000 мкм2. Швидкість кровотоку в капілярі порівняно невелика — 0,3 мм/с, що дозволяє еритроцитові перебувати в капілярі близько 2—3 с. Залежно від будови стінки розрізняють капіляри соматичного, вісцерального та синусоїдного типу.
Стінка капіляра складається з одного шару ендотеліальних клітин. Ендотеліоцити в капілярах, як і в інших відділах судинного русла, є активними елементами судинної стінки. В них синтезуються різні ферменти і біологічно активні сполуки, наприклад, антитромбін III, ендотеліальний фактор розслаблення (ЕФР) і скорочення (ЕФС), які активізують або гальмують дію гормонів, медіаторів на судинну стінку, факторів тромбоутворення або ж самі спричиняють фізіологічні ефекти.
Доведено, що за деяких умов ендотеліоцити можуть скорочуватися і з плоских ставати об'ємнішими. В них містяться мікро-фібрили, що складаються з актинових, міозинових та інших скоротливих елементів. Ці утворення розташовані переважно вздовж основи клітин і прикріплюються до цитолеми в місцях міжклітинних контактів (мал. 163). Скорочення мікрофібрил може спричинити два ефекти: розходження клітин і збільшення міжклітинної щілини або ж зміни форми клітини і випинання її в просвіт судини. Конкретний наслідок визначається тиском крові на стінку капіляра: якщо він досить великий, то закривається просвіт капіляра
Регуляція кровотоку
в мікроциркуляторному руслі
У зв'язку з відсутністю в стінках капілярів гладком'язових волокон, які могли б активно змінити їх просвіт, функція регуляції кровотоку через них належить артеріолам, метартеріолам, прекапі-лярним сфінктерам, посткапілярним венулам та артеріовенулярним шунтам. Кінцеві артерії та артеріоли містять у стінці відносно велику кількість гладком'язових волокон. У міру зменшення діаметру артерій товшина М'язового шару поступово зменшується, і в артеріолах м'язові клітини розташовані вже в один шар. У місцях переходу артеріол у капіляри м'язові клітини зустрічаються все рідше.
Лекція № 7.
Тема: Фізіологія спинного мозку і стовбура мозку.
План:
Будова спинного мозку.
Висхідні та низхідні шляхи спинного мозку. Пірамідний шлях.
Спинномозкові рефлекси: розтягнення, ритмічні, тонічні, шкірні.
Спінальний шок, видові особливості.
Будова довгастого мозку та мосту. Черепно-мозкові нерви.
Задній мозок: тонічні рефлекси статичні та статокінетичні, тонічні лабіринтні та шийні рефлекси.
Середній мозок, будова. Децеребральна ригідність. Статичні, статокінетичні, орієнтовні та сторожеві рефлекси.
Ядра таламуса та гіпоталамуса в регуляції організму.
Роль ретикулярної формації в регуляції функцій організму.
1. Спинний мозок регулює рухи та діяльність вегетативних структур тулуба. Він має сегментарну будову (31 сегмент). Сегменти складають відділи: шийний, грудний, поперековий, крижовий, куприковий. На поперечному розрізі СМ має сіру речовину, яка нагадує метелика з виступами назад, вперед і в боки (роги). Сіра речовина складається з тіл нейронів та їхніх відростків. Навколо сірої розташована біла речовина (канатики), яка побудована нервовими волокнами, що утворюють висхідні і низхідні шляхи. Від кожного сегмента СМ вправо і вліво відходять по парі корінців – задніх і передніх. Задні корінці побудовані з пучків чутливих нервових волокон, а передні утворені руховими нервовими волокнами. Це легко довести дослідом на жабі, поперемінно перерізуючи корінці, подразнюючи їх і реєструючи струми дії при скороченні лапок. На деякій відстані від білої речовини корінці зливаються, утворюючи змішаний нерв. У місці їх зустрічі знаходиться потовщення, де розташовані чутливі нейрони. Корінці кожного сегмента іннервують ділянку тіла, яка називається метамером.
Розрізняють дві функції СМ – рефлекторну і провідникову.
2. Шляхи СМ проходять по канатикам і поділяються на чутливі (висхідні) і
рухові (низхідні).
А) Передній канатик – утворений нейритами (аксонами), що передають нервові імпульси від вищих центрів до нижчих і формують такі нервові шляхи: передній пірамідний =кірково-спинномозковий (регулює свідомі рухи), передвірно-спинномозковий (координує рухи і орієнтацію тіла в просторі), передній спинно-таламічний (дотик).
Б) Бічний канатик – складається з аксонів, що проводять імпульси в двох напрямках – від вищих центрів до нижчих і навпаки: бічний пірамідний, червоно ядерно-спинномозковаий або екстрапірамідний (несвідомих рухів), бічний спинно-таламічний (шлях болі, температури).
В) Задній канатик – проходять відростки чутливих клітин спинномозкових вузлів, що утворюють тонкий шлях Голя і клиноподібний шлях Бурдах ( (несуть сигнал від пропріоцептивних подразнень в ядра проміжного мозку).
Отже, основними висхідними шляхами є :
- шлях Голя і Бурдаха (усвідомлена пропріоцептивна чутливість),
- задній і передній спинно-мозочковий шлях (передають пропріоцептивну інформацію в мозочок),
- бічний спинно-таламічний (тактильна, больова і температурна чутливість).
Основними низхідними шляхами є:
- бічний і передній пірамідний (свідомі рухи),
- екстрапірамідний (несвідомих рухів і тонусу м’язів),
- перед вірно-спинномозковий і сітчасто-спинномозковий (несвідомі рухові реакції і тонус м’язів на збереження рівноваги і координації рухів).
3. Рефлекторна діяльність СМ зв’язана з рефлекторними дугами, які замикаються на рівні нервових центрів СМ. Для вивчення функцій СМ його відділяють від головного. Тварина без головного мозку називається спінальною. На спінальних тваринах вдалося з’ясувати, що у нижчих хребетних СМ забезпечує зберігання пози, локомоцію (ходьба), захисні і статеві рефлекси. У вищих хребетних у зв’язку з розвитком головного мозку СМ повністю забезпечити їхні фізіологічні функції не може.
Рефлекси СМ поділяються на рухові (тонічні і фазні) і вегетативні (регулюють роботу внутрішніх органів, судин, залоз).
І. Рухові рефлекси.
А) Тонічні (полісинаптичні)– регуляція СМ м’язового тонусу, це необхідно для збереження положення тіла (н-д, нахил голови при читанні), підтримання пози при ходьбі. В основі цих рефлексів лежить пропріоцептивна дія ( від мотонейронів в нечастому ритмі імпульси йдуть до м’язів і підтримують постійну напругу м’яза), а також тонічні рефлекторні впливи з боку вищих відділів ЦНС.
Б) Фазні (швидкі) рефлекси- виникають при подразненні різних рецепторів. Їх можна спостерігати при нанесенні на шкіру спінальної жаби механічних та хімічних подразнень, при цьому можна отримати більшу кількість рефлекторних фазних реакцій (згинання, почухування, крокування та ін.) - сухожилкові, шкірні, ритмічні=локомотивні рефлекси.
Рефлекторна діяльність СМ корегується вищими центрами, які знаходяться в головному мозку. Морфологічною основою цієї діяльності є висхідні і низхідні шляхи, які зв’язують нервові центри СМ з головним і навпаки.
1) Рефлекси розтягнення (сухожилкові) – при незначному розтягуванні ( всього на 0,1 мм) у рецепторах м’яза виникає збудження, яке рефлекторно викликає скорочення. Аналогічні рецептори є і в сухожилках. Оскільки сухожильні рефлекси на розтягнення виникають у відповідь на подразнення рецепторів м’яза, їх називають власними рецепторами м’язів (пропріорецепторами). Н-д, колінний сухожильний (моносинаптичний) рефлекс.
2) Ритмічні рефлекси – це група рефлексів поперемінної дії згиначів і розгиначів кінцівок. Виникає при тривалій дії несильного подразника. За допомогою цих рефлексів здійснюється ходьба і біг. Якщо спінальній собаці нанести подразнення стопи уколом, його кінцівки починають ритмічно рухатися (крокування як при ходьбі). Тобто на рівні спинного мозку розміщений «локомотивний» центр ходьби. Прикладом ритмічних рефлексів є почухувальний рефлекс
3) Шкірні рефлекси (згинальні, полісинаптичні) виникають при подразненні рецепторів шкіри. Н-д, при подразненні шкіри стопи спостерігається її підошовне згинання (підошовний рефлекс).
ІІ. Вегетативні рефлекси – вісцеро-вісцеральні (н-д, зміна тонусу судин при зміни тиску в аорті і каротидному синусі), вісцеро - дермальні рефлекси (н-д, подразнення внутрішніх органів і зміна шкірної чутливості на визначеної ділянці), дермато-вісцеральні (н-д, подразнення шкіри викликають судинні реакції), сомато-вісцеральні ( н-д, скорочення м’язів викликає зміну роботи внутрішніх органів), вісцеро-соматичні.
4. Спінальний шок – результат припинення впливів вищих відділів ЦНС на спинний мозок. Проявляється тимчасовим припиненням рефлекторної діяльності спинного мозку. У спінальної жаби (спінальна жаба – жаба, у якої зруйнований головний мозок) явище спінального шоку продовжується декілька хвилин, у кішки – декілька днів, у приматів – декілька неділь. У приматів після відновлення рефлекторної діяльності СМ частина рефлексів змінюється в порівнянні з інтактними тваринами, а частина не відновлюється зовсім. Ведучим механізмом в розвитку спінального шоку є припинення активуючих впливів структур головного мозку на нейрони СМ. Чим вищий рівень організації тварини, тим більш важливими є ці впливи.
5. Довгастий мозок – це продовження спинного мозку. Передня поверхня розділена передньою серединною щілиною, з боків якої розташовані два повздовжніх підвищення – піраміди, які утворені пучками нервових волокон пірамідних провідних шляхів, які з’єднують кору великого мозку з ядрами черепних нервів і передніми рогами спинного мозку. На задній поверхні проходять тонкий і клиноподібний канатики, які вміщують нервові шляхи. Вони з’єднують спинний мозок з однойменними ядрами довгастого мозку. Латерально від пірамід лежать оліви. Крім того, в склад сірої речовини довгастого мозку входять ядра 9-12 пар черепних нервів і ретикулярна формація. Довгастий мозок виконую провідну і рефлекторну функцію (центри захисних рефлексів, центри дихання, кровообігу, травлення, установочні рефлекси – забезпечують тонус м’язів для підтримки пози).
Задній мозок утворений мостом і мозочком. Міст (Варолієв міст) розташований вище від довгастого мозку і нагадує потовщений валик. Задня поверхня моста бере участь в утворенні ромбоподібної ямки. Сіра речовина моста утворена власними ядрами моста, ядрами черепних нервів ( 5-8 пари) і ядрами ретикулярної формації. Біла речовина складається з нервових волокон, які утворюють провідні шляхи, які на передній поверхні йдуть в довгастий мозок і подальше в спинний мозок. В задній частині волокна утворюють висхідні і низхідні шляхи двобічних зв’язків мосту з вище розташованими структурами і мозочком.
Черепні нерви. Від стовбура головного мозку відходить 12 пар черепних нервів. Їх ядра розташовані в сірої речовині головного мозку. Вони поділяються на чутливі, рухові та змішані (чутливо-рухові) нерви. Чутливі нерви – І пара (нюховий нерв), ІІ пара (зоровий нерв), YІІІ пара (слуховий нерв), рухові нерви - іннервують м’язи ока (ІY пара – блоковий нерв, , YІ пара – відвідний нерв), м’язи шиї і язика (ХІ пара – додатковий нерв і ХІІ пара – під’язиковий нерв), змішані нерви – іннервують м’язи ока і м’яз-звужувач зіниці ( ІІІ пара – окоруховий нерв), мімічні м’язи і слинні залози (YІІ пара – лицевий нерв), смакові сосочкі язика, піднебінні мигдалини, біля вушну слинну залозу (ІХ пара – язикоглотковий нерв), жувальні м’язи, шкіру обличчя, зуби, слинні залози, слизову оболонку рота (Yпара – трійчастий нерв) , слизову оболонку і м’язи внутрішніх органів (Х пара – блукаючий нерв і ХІ пара – додатковий нерв).
Рухові функції стовбура головного мозку.
Найбільш важливими руховими центрами стовбура мозку є: латеральне вестибулярне ядро моста, червоне ядро середнього мозку, деякі ядра ретикулярної формації. Стовбурові центри формують тонічні рефлекси (забезпечують м’язовий тонус), які поділяються на дві групи:
Статичні рефлекси – збереження рівноваги і нормального вертикального положення тіла:
А) позові або рефлекси положення тіла в просторі – лабіринтні (=вестибулярні) і шийні;
Б) випрямні (установні).
Статокінетичні рефлекси – рух тіла в просторі:
А) рефлекси на обертові прискорення – подразнення рецепторів півколових каналів;
Б) рефлекси на прямолінійне прискорення – подразнення рецепторів передвір’я.
Статичні рефлекси виникають при зміні положення тіла, особливо голови в просторі. Так, наприклад, прояв шийних рефлексів – нахил голови вперед підвищує тонус всіх м’язів-згиначів, а назад – розгиначів. Вестибулярні рефлекси - це підвищення або зниження тонусу м’язів усіх кінцівок, які направлені на компенсацію відповідного руху голови для попередження можливого падіння під час переміщення тіла і голови в просторі.
Статичні випрямні рефлекси пов’язані, насамперед, з подразненням вестибулярних рецепторів при положенні голови униз. При цьому запускаються ланцюги послідовних рухів, які направлені на відновлення указаної пози (при подразненні пропріорецепторів м’язів шиї, рецепторів поверхні тіла, зору). Наприклад, ці рефлекси проявляються під час навчання плаванню. Людина, яка поглинає у воду, пробує зайняти горизонтальне положення: підіймається голова, вслід за цим підіймається і тулуб (людина встає на дно).
Статокінетичні рефлекси проявляються під час переміщення тіла в просторі, при зміні швидкості руху. Наприклад, «ліфтові» рефлекси (подразнення рецепторів передвір’я) - на початку руху вгору збільшується тонус м’язів – згиначів, а зупинка призводить до збудження розгиначів.
Отже, тонічні рефлекси забезпечують перерозподіл тонусу м’язів між різними групами м’язів, таким чином попереджається можливість порушень рівноваги і втрата активної пози.
6. Середній мозок. Середній мозок складається з чотиригорбистості і ніжок мозку. Від чотиригорбистості відходять ручки до проміжного мозку (до медіальних і латеральних колінчастих тіл). На розрізі ніжка мозку складається з покришки і основи, межею між ними є чорна речовина.
Функції середнього мозку:
1) Провідникові - в основі проходять низхідні нервові шляхи від кори в спинний мозок, в покришці – висхідні шляхи до таламусу.
2) Рефлекторні – в покришці є червоне ядро (починає несвідомий руховий шлях) і ретикулярна формація.
Важливі рухові ядра СрМ:
Червоні ядра – починаються руброспінальні шляхи.
Ретикулярна формація.
Чорна речовина – функціонує як єдине ціле з базальними ядрами.
За участю рухових ядер СрМ здійснюється такі рефлекси:
1) Випрямні (статичні)– забезпечують відновлення порушеної пози. У тварин, що мають шию, випрямні рефлекси мають 2 фази:
А) Рефлекс випрямлення голови – він починається з подразнення отолітових вестибулорецепторів у відповідь на порушення нормального положення голови в просторі ( наприклад, при падінні). Рівень замикання – червоні ядра, РФ. По низхідним шляхам від цих структур інформація передається до рухових нейронів СМ, йде перерозподіл тонусу м’язів шиї, голова набуває нормальне положення в просторі (тім’ячком доверху).
Б) Рефлекс випрямлення тулуба – починається з подразнення пропріорецепторів шиї, що спричинюється рефлексом випрямлення голови, так як при цьому відновлюється нормальне положення голови у просторі, але порушується її нормальне положення відносно тулуба. Вхід інформації від пропріорецепторів шиї здійснюється на рівні СМ, підйом до структур СрМ, обробка цієї інформації, передача до мотонейронів СМ, до м’язів, які забезпечують відновлення нормального положення тулуба відносно голови в просторі.
2) Статокінетичні – забезпечують збереження пози рівноваги при русі з прискоренням. При русі з лінійним прискоренням збуджуються отолітові рецептори, а при русі з кутовим прискоренням – рецептори півколових канальців. Вхід цієї інформації в ЦНС здійснюється на рівні заднього мозку, підйом до структур СрМ, обробка цієї інформації, передача низхідними шляхами до мотонейронів СМ, перерозподіл тонусу м’язів, який забезпечує збереження рівноваги під час руху з прискоренням. Важливим компонентом цих рефлексів є присінкові-окорухові рефлекси, ністагм очей, більш продовжена фіксація предметів в полі зору під час руху, краща орієнтація в просторі, більш надійне збереження пози при русі.
Ці рефлекси неможливо отримати у децереброваної чи спінальної тварини.
За рахунок обробки інформації від зорових та слухових рецепторів рухові ядра СрМ забезпечують здійснення орієнтацій них рефлексів – у відповідь на сильні звукові та світлові подразники тварина повертає голову в бік подразника, насторожує вуха, в неї дещо посилюється тонус флексорів – тварина ніби готується до відповіді на цей подразник (стартове положення до бігу або захисту).
Вивчити роль заднього мозку в регуляції рухових функцій можна на тваринах з децеребрацією – перерізкою стовбура мозку між середнім та заднім мозком (точніше – нижче червоних ядер середнього мозку). Після операції децеребрації у тварини виникає децеребральна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті, голова закинута назад за рахунок розгинання шиї; хвіст піднятий, тулуб розігнутий. Ця поза пов’язана з гіпертонусом розгиначів. В основі розвитку
децеребральної ригідності лежать вестибулоспинальні впливи, завдяки яким відбувається: посилення тонічних рефлексів екстензорів і послаблення тонічних рефлексів флексорів.
7. Ядра таламуса і гіпоталамуса в регуляції організму.
Таламус є структурою проміжного мозку і виконує сенсорні функції – за виключенням нюхового, в таламусі переключаються всі сенсорні шляхи, які йдуть в кору ГМ. В таламусі є біля 40 ядер, які розрізняються за своїми функціями. Серед них виділяють:
Специфічні (сенсорні) ядра – отримують інформацію від специфічних сенсорних шляхів, переробляють її і передають в сенсорні зони КГМ, де виникають відповідні відчуття (зорові, слухові та ін..).
Неспецифічні ядра – отримують інформацію від ретикулярної формації стовбура мозку по шляхах больової чутливості. Вони передають інформацію до всіх зон КГМ, здійснюючи на неї неспецифічний активуючий вплив (підвищують збудливість різних ділянок кори).
Асоціативні ядра – отримують інформацію від специфічних сенсорних перемикаючих ядер і від неспецифічних ядер таламуса. Вони передають інформацію в асоціативні зони кори ГМ (лобова, тім’яна, скронева). Разом з даними ділянками кори, асоціативні ядра таламуса приймають участь в розпізнаванні образів.
Неспецифічні перемикаючі ядра – лімбічні (передають інформацію від гіпоталамуса в лімбічну кору – формуються емоції і мотивації) і рухові (передають інформацію від базальних ядер, мозочка в рухову кору – забезпечення рухових реакцій).
При ураженні таламуса у людини виникає повна втрата чутливості або її зниження на протилежному боці, випадає скорочення мімічних м’язів, яке супроводжує емоції, також можуть виникати порушення сну, зниження слуху, зору та ін..
Гіпоталамічна ділянка бере участь в регуляції різних видів обміну речовин, регуляцію теплоутворення і тепловіддачу, стан сну і бадьорості. Також утворюються гормони, які потім депонують в задньої частці гіпофіза. Гіпоталамічна ділянка регулює вегетативні функції: передні відділи гіпоталамуса є вищі центри парасимпатичної НС, задні – симпатичної НС.
8. Ретикулярна формація (РФ) – скупчення нервових клітин, дендрити та аксони їх формують низхідні та висхідні шляхи сітчастої речовини.
Висхідні зв’язки:
Через неспецифічні ядра таламуса РФ передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини: забезпечується бадьорий стан організму та визначається рівень активності кори. РФ має окремі структури, котрі зменшують рівень активності кори і зумовлюють стан сну (гіпогенні зони).
РФ передає інформацію від різних рецепторів в гіпоталамус.
Низхідні зв’язки:
РФ специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок (ретикулоспинальні шляхи).
Деякі шляхи неспецифічно активують чи гальмують спин альні мотонейрони і визначають ступень їх активності.
Від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспинальним шляхам передається інформація до симпатичних рухових нейронів СМ при регуляції відповідних функцій організму.
Лекція №9.
Тема: Фізіологія дихання.
План:
І. Будова і функції системи дихання.
ІІ. Етапи процесу дихання. Дихальний цикл. Вентиляція легень.
ІІІ. Опір диханню. Склад повітря, що вдихається і видихається.
ІУ. Статичні і динамічні показники функції зовнішнього дихання.
У. Транспорт кисню кров’ю. Крива дисоціації оксигемоглобіну.
Киснева ємність крові.
УІ. Газообмін між кров’ю та тканинами.
І. Дихальний апарат складається з повітроносних шляхів та легень. Повітроносні шляхи починаються верхніми дихальними шляхами, до яких відносять: носові ходи, глотка, гортань. Далі повітря потрапляє в трахею, де поділяється на два головних бронхи, які поділяються на часткові і сегментарні бронхи та дихальні бронхіоли. Бронхіола продовжується альвеолярними ходами, переходячи у сліпі мішечки – альвеоли. Стінка бронхів і бронхіол непроникна для газів, але тут повітря очищується, зігрівається, зволожується. Тому з функціональної точки зору цю повітроносну зону називають мертвим простором, де повітря очищується, зволожується і зігрівається. Його об’єм у чоловіків дорівнює 150 мл, у жінок – 100 мл. Слизова оболонка дихальних шляхів вкрита епітелієм, серед клітин якого розрізняють клітини війчасті та секреторні, що продукують слиз. У товщі слизової оболонки розташована густа капілярна сітка. Повітря, перед тим як потрапити до легенів, нагрівається до 37 градусів, зволожується і очищається від твердих частинок. Війки епітелію ритмічно, швидко рухаються у напрямку до глотки і повільно – в зворотному напрямку. Завдяки цим рухам слиз з налиплими на ньому часточками проштовхується до глотки зі швидкістю, більшою за 1 см/хв., де відкашлюється або ковтається. Дихальну зону легень складають ацинуси, до яких відносять дихальні бронхіоли, альвеолярні ходи та альвеоли. Кожна альвеола оточена густою сіткою кровоносних капілярів.
ІІ. Диханням називається сукупність процесів, що забезпечують потреби організму у кисні і виділенні вуглекислого газу. Надходження кисню необхідне для окислення органічних речовин, в результаті чого звільняється енергія, що потрібна для життєдіяльності організму. Вуглекислий газ, як кінцевий продукт, повинен бути видалений з організму. Крім газообмінної функції, дихання сприяє підтриманню найважливішої константи внутрішнього середовища – рН рідин і тканин. Так, при зсуві рН в кислу сторону, за допомогою прискорення дихання виводиться більше вуглекислого газу. Дихання також бере участь у підтриманні температури тіла на постійному рівні.
Дихання складається з наступних процесів:
І етап. 1) Зовнішнє дихання або вентиляція легень – обмін газів між організмом і
зовнішнім середовищем;
2) Обмін газів у легенях - відбувається між альвеолярним повітрям і кров’ю
капілярів малого кола кровообігу;
ІІ етап. Транспорт газів – перенос кров’ю кисню від легень до тканин і вуглекислого газу
від тканин до легень;
ІІІ етап. 1) Обмін газів між кров’ю великого кола кровообігу і клітинами тканин;
2) Внутрішнє або тканинне дихання – це біологічне окислення в мітохондріях
клітин.
Дихальний цикл – це вдих (inspiracio)= 0,9-4,7 с, видих (exspiracio)= 1,2-6 с, та дихальна пауза (непостійна, навіть може бути відсутньою).
Механізм вдиху:
1.Скорочення зовнішніх міжреберних м’язів – підіймання ребер, груднина рухається вперед, об’єм грудної клітки збільшується в фронтальній і сагітальній площині.
2. Скорочення діафрагми – купол стає плоским, грудна порожнина збільшується в вертикальному напрямку.
3.Скорочення допоміжних дихальних м’язів грудної клітки і живота – під час глибокого вдиху.
Механізм видиху:
1.Розслаблення міжреберних м’язів – грудна клітка під силою тяжіння повертається у вихідне положення.
2. Розслаблення діафрагми – підіймається купол, зменшується об’єм грудної клітки.
3. Скорочення внутрішніх міжреберних м’язів, м’язів черевного пресу – при форсованому видиху.
При кожному дихальному циклі повітря у легенях оновлюється. Цей процес називають вентиляція легень. Кількісним показником її є хвилинний об’єм дихання (ХОД). ХОД – кількість повітря, що вдихається і видихається протягом однієї хвилини. ХОД=ДО х ЧД за 1 хв. ЧД залежить від фізичного навантаження. У стані спокою ЧД =12-24 дих.рухи, що забезпечує легеневу вентиляцію 6-10 л/хв.. При фізичному навантаженні ХОД збільшується в 5-10 разів.
Так, якщо при спокійному диханні в легені надходить близько 500 мл повітря, то цей об’єм додається до ЗО і РОвид, який дорівнює приблизно 2000 мл. Але частина повітря, що видихається не доходить до альвеол і залишається в дихальних шляхах.
У зв’язку з наявністю МП альвеолярна вентиляція відрізняється від легеневої: із 500 мл повітря до альвеол не доходять 150 мл. Тобто за кожний дихальний цикл до альвеол надходить близько 350 мл повітря, що складає приблизно 1/7 всього повітря, що міститься в альвеолах. Природно, що чим глибше дихання, тим інтенсивніша альвеолярна вентиляція, оскільки з одного боку, при глибшому видиху в легенях залишається менше повітря, а з другого – при форсованому диханні істотно збільшується ДО.
Для характеристики дихання людини визначають ще ряд динамічних показників, що дають уявлення про дихання за певний час (частіше за 1 хв). До них належать такі.
Частота дихальних рухів (ЧД).
Хвилинний об’єм дихання (ХОД) – кількість повітря, що надходить у легені за 1 хв
Альвеолярна вентиляція (АВ) характеризує вентиляцію альвеол:
АВ= (ДО-МП) х ЧД.
Максимальна вентиляція легень (МВЛ) – кількість повітря, яке людина вдихає і видихає при максимальній глибині і частоті дихання.
Резерв дихання – різниця МВЛ і ХОД.
Коефіцієнт легеневої вентиляції (КЛВ) – та частина повітря, яка обмінюеться в легенях під час кожного вдиху: КЛД=(ДО-МП):ФЗЄ.
Коефіцієнт альвеолярна вентиляція/легеневий кровотік: АВ:ЛК=4 л:5л=0,8.
ІІІ. Опір диханню . Сурфактант.
Види опору диханню:
Нееластичний опір (динамічний, в’язкий опір) –тертя тканин дихального апарату, опор потоку повітря у повітроносних шляхах, зумовлений тертям частин повітря між собою і між поверхнею провідних шляхів. Цей опір залежить від багатьох факторів – у здорової людини від об’єму дихання, у хронічного курця – від посилення секреції слизу в дихальних шляхах і звуження бронхів веде до постійного підвищення опору дихання.
Еластичний опір – опір легеневої тканини і грудної клітки. Він складається з еластичної тяги легеневої тканини і всього дихального апарату, а також з поверхневого натягу вологи альвеолярних стінок. Для зменшення останнього клітини альвеолярного епітелію декретують спеціальну речовину – сурфактант (ліполпротеїд), що утворює на поверхні альвеол плівку. Молекула сурфактанту однойменно заряджена і відштовхують один від одного , перешкоджаючи злипанню альвеол під час видиху. Крім того, сурфактанти беруть участь в очищенні альвеол від пилових часток, сприяють збереженню сухості альвеол, посилюють фагоцитарну активність легеневих макрофагів.
Еластичний опір легенів бере участь в побудові еластичної тяги легень. Еластична тяга легень – здатність легень стискатися після їх розтягнення (властивість гуми).
Фактори еластичної тяги:
Поверхневий натяг альвеол – альвеоли прагнуть спадаться при їх розтягненні.
Постійний тонус м’язів бронхіального дерева
Еластичні волокна в тканинах легень.
Еластична тяга легень визначает величину тиску в плевральній порожнині – внутрішньоплевральний тиск. Він нижче атмосферного (негативний) на величину еластичної тяги. Величина внутрішньо плеврального тиску залежить від фаз дихальних рухів. При видиху він досягає -3 мм рт.ст., при видиху – 6 мм рт.ст. Завдяки цьому легені знаходяться в розтягнутому стані. В випадку потрапляння повітря в плевральну порожнину еластична напруга легенів призводить до їх спадання - пневмотораксу.
Газообмін в легенях. В період вдиху повітря струйно доходить лише до 17 генерації бронхіол. Починаючи з них, до струйного потрапляння повітря приєднується дифузний засіб обміну кисню і вуглекислого газу. При дифузії руховою силою газообміну є різниця парціальних тисків газів між повітроносними шляхами і альвеолами. За рахунок цього кисень дифундує в альвеоли, а в протилежному напрямку йде вуглекислий газ.
Склад повітря.
Склад повітря, що вдихаємо: О2 = 20,9%, СО2 =0,03%, азот =79%.
Склад повітря, що видихаємо: О2=16%, СО2=4,5%, азот = 79,5%.
Альвеолярна газова суміш: О2=14%, СО2=5,5%, азот = 80,5%.
Альвеолярне повітря є внутрішнім газовим середовищем. Його постійність зумовлює нормальний газовий склад артеріальної крові.
ІУ. Статичні і динамічні показники функції зовнішнього дихання.
Ці показники залежать від розмірів грудної клітки та її рухомості.
А. Статичні показники.
Дихальний об’єм (ДО) – кількість повітря, яке потрапляє в легені за один спокійний вдих. ДО=500 мл.
Резервний об’єм вдиху (РОвд) – максимальна кількість повітря, яке людина може вдихнути після нормального вдиху (2500 мл)
Резервний об’єм видиху (РО вид) – максимальна кількість повітря, яке людина може видихнути після спокійного видиху (близько 1500 мл).
Життєва ємкість легень (ЖЕЛ) –найбільша кількість повітря, яке людина може видихнути після максимального глибокого вдиху. ЖЕЛ = ДО + РОвд+РОвид. ЖЄЛ залежить від росту, маси тіла і фізичного розвитку.
Залишковий об’єм (ЗО) повітря, що залишається після максимального глибокого видиху в легенях. ЗО= 1л.
Загальна ємкість легень (ЗЕЛ) – кількість повітря, яке знаходиться в легенях на висоті максимального вдиху: ЗЕЛ=ЖЄЛ=ЗО
Об’єм дихальних шляхів (мертвий простір) – 150 мл.
Функціональна залишкова ємкість (ФЗЄ) – кількість повітря, яке залишається в легенях в кінці видиху. ФЗЄ= РОвид +ЗО.
Б. Динамічні показники – відображають ефективність функціонування системи дихання в часовому аспекті (зазвичай за 1 хв:
1. Частота дихальних рухів за 1 хвилину = ЧД
2. Хвилинний об’єм дихання (ХОД) – кількість повітря, яке проходить через легені за 1 хвилину. ХОД=ДОхЧД.
3.Альвеолярна хвилинна вентиляція (АВ) характеризує вентиляцію альвеол:
АВ=(ДО-МП)хЧД
4. Максимальна вентиляція легень – кількість повітря, яке можна вдихнути при максимальній частоті і глибині дихання.
У. Транспорт дихальних газів кров’ю. Крива дисоціації оксигемоглобіну.
Киснева ємність крові.
Кисень у крові знаходиться у двох станах: фізичному розчиненні (незначно) і в хімічному зв’язку з Нв. Нв утворює з киснем нестійке, легко дисоційоване з’єднання – оксигемоглобін. Кожна молекула Нв здатна приєднати 4 молекули кисню, що в перерахунку на 1 грам Нв - 1,34 мл О2.
Кількість кисню, що може окислити 100 мл крові при повному насиченні гемоглобіном, називають кисневою ємкістю крові (КЄК)(= кількість О2, яке зв’язується кров’ю до повного насичення Нв; або - максимальна кількість кисню, яке може бути зв’язаним 100 мл крові).
КЄК= Нв х1,34
Беручи до уваги, що в 100 мл крові вміщується лише 0,3 мл розчиненого О 2 можна зробити висновок, що основна кількість транспортуємого кров’ю кисню – у вигляді хімічного зв’язку з гемоглобіном.
При середній концентрації гемоглобіну (15%) у чоловіків КЄК дорівнює 20 мл, у жінок – 18 мл О2 /100 мл крові. При фізичних навантаженнях КЄК збільшується до 24%, тому що в циркулюючу кров виходять еритроцити з депо.
Насичення Нв киснем близько 97%. Ступень насичення Нв киснем і дисоціація оксиНв (утворення відновленого Нв) не знаходять в прямій пропорційній залежності від напруження кисню (парціального тиску). Інтенсивність утворення оксигемоглобіну обумовлена парціальним тиском О2 в крові: чим вище рівень Р О2, тим більше утворюється НвО2. Це не прямо пропорційна залежність у графічному виразі називають кривою дисоціації оксигемоглобіну. Крива дисоціації оксигемоглобіну має S-подібну форму.
При аналізі цієї залежності можна зазначити, що зменшення парціального тиску кисню у крові спричинює дисоціацію оксигемоглобіну – звільнення гемоглобіну від О2(відновлений Нв). Відсотковий вміст оксигемоглобіну зменшується, а відновленого – збільшується. Отже, при зниженні парціального тиску кисню в крові ступінь насичення гемоглобіну киснем зменшується. що відображає властивості гемоглобіну, який майже повністю насичується киснем при значних коливаннях його парціального тиску в альвеолярному повітрі. Дихання чистим киснем може збільшити оксигенацію крові тільки на 3%. Таким чином, плоска верхня частина S-подібної кривої дисоціації оксигемоглобіну свідчить про те, що організм має надійний захист від значного зниження парціального тиску кисню в альвеолах. Крута частина кривої показує, що при зниженні тиску кисню (у капілярах тканин) дисоціація оксигемоглобіну відбувається дуже швидко, і тканини мають надійний «захист» від нестачі кисню (гіпоксії). Швидкість дисоціації оксигемоглобіну збільшується при зниженні рН крові, підвищенні її температури і збільшенні кількості СО2.
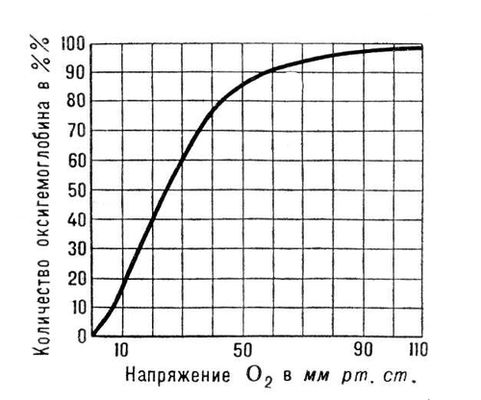
При відсутності кисню у крові (РО2=0) оксигемоглобіну в крові немає, а при РО2 =10 мм рт.ст. 10% Нв переходить в оксигемоглобін, то при РО2=20 мм рт.ст. НвО2 вже складає 30%. Максимальна кількість Нв (45-80%) зв’язується з киснем при його напруженні 30- 40 мм рт.ст. (утворюється біля 80% НвО2). Подальше підвищення напруження кисню приводить до зниження швидкості утворення окси Нв.
Спорідненість Нв до кисню значно знижується при зрушенні реакції крові в кислий бік (ацидоз), що спостерігається в тканинах організму внаслідок утворення СО2. Ця властивість Нв має важливе значення для організму. В капілярах тканин, де концентрація СО2 в крові збільшена, здатність Нв утримувати О2 зменшується, що полегшує його віддачу клітинам. В альвеолах легень, де частина СО2 переходить в альвеолярне повітря, здатність Нв зв’язувати кисень знову зростає.
Перехід Нв в оксиНв та з нього в відновлений залежить від температури. При одному ж тому тиску О2 в оточуючому середовищі при температурі тіла 37-38 градусів найбільша кількість оксиНв переходить в відновлений Нв.
Отже:
Транспорт О2 забезпечується в основному за рахунок зв’язку з Нв.
Насиченість Нв киснем залежить в першу чергу від парціального тиску газу в атмосферному і альвеолярному повітрі.
Основна причина віддачі кисню гемоглобіном є зсув реакції в тканинах в кислий бік.
Транспорт вуглекислого газу відбувається фізичним (розчинення) і хімічним шляхами. Розчинений у плазмі крові СО2 становить близько 5% загальної кількості вуглекислого газу, що транспортується; 95% його переноситься у хімічно зв’язаному стані. Спрощено процес транспорту СО2 можна уявити так: СО2 потрапляючи у кров, приєднує воду, внаслідок чого утворюється вугільна кислота, яка відразу з’єднується з білками плазми:
Nа (білка) +Н2СО3 =NаНСО3+Н (білка). У легенях NаНСО3 дисоціює, і звільнений СО2 виходить в альвеоли. Процес утворення вугільної кислоти і її дисоціація прискорюються ферментом карбоангідразою, який міститься в еритроцитах. Залежно від напруження СО2 карбоангідраза каталізує утворення вугільної кислоти і її розщеплення на СО2 і воду (у капілярах легень). Частина молекули СО2 сполучається в еритроцитах з гемоглобіном, утворюючи карбгемоглобін НвСО2 . У капілярах тканин гемоглобін віддає кисень, чим полегшує насичення крові СО2.
УІ. Газообмін між кров’ю та тканинами.
Кисень – дифузія кисню з крові в тканини здійснюється за його градієнтом. Внаслідок різниці тиску О2 на межі капіляра і клітини (100 мм рт.ст.) відбувається дисоціація оксиНв і рух розчиненого кисню через стінку капіляра, міжклітинний простір, внутрішньоклітинний простір, оболонку мітохондрій (в мітохондріях кисень використовується для хімічних реакцій окислення). Якщо потреби клітини в енергії зростають, і тиск кисню біля мітохондрій падає нижче критичного рівня, тоді О2 для окислення не вистачає, і клітина переходить на анаеробний шлях енергозабезпечення з утворенням молочної кислоти. Не весь кисень, який потрапляє в капіляри, використовується тканинами. Частина його залишається у венозній крові. Різницю між відсотковим вмістом О2 в артеріальній і венозній крові називають коефіцієнтом тканинної утилізації кисню (КТУК). У стані спокою він дорівнює 25-30%. Це означає, що тканини за час перебування порції крові в капілярі встигають поглинути лише ¼ кисню, що до них надходить. При напруженій м’язовій роботі КТУК зростає до 50-60%. Під час напруженої діяльності, коли м’язові волокна стискають капіляри, де утрудняється і навіть припиняється кровообіг, у постачанні киснем міокарда і скелетних м’язів бере участь міоглобін. Його особливістю є те, що вже при РО2=10 мм рт.ст., він повністю переходить у міооксигемоглобін. Зв’язок О2 із залізом у міоглобіні міцніший, тому міоглобін віддає кисень тільки при гіпоксії.
Вуглекислий газ переходить з тканин у кров завдяки різниці його парціального тиску. У мітохондріях РСО2 становить 60 мм рт.ст., у тканиній рідині – 48 мм рт.ст., у венозній крові – 46 мм рт.ст.
Лекція №10.Тема: Регуляція дихання.План:
І. Структури ЦНС, що регулюють ритм дихання: дихальний, пневмотаксичний, апнейстичний центри заднього мозку.
ІІ.Роль центральних і периферичних рецепторів у забезпеченні газового гомеостазу.
ІІІ. Дихання під час фізичної роботи, при підвищенному та заниженому барометричному тиску.
ІУ. Механізм першого вдиху новонародженого.
Дихальний центр розташований в ділянці стовбура мозку, в довгастому мозку.
В довгастому мозку є скупчення нейронів, які проявляють свою активність під час вдиху або видиху – дорсальні і вентральні ядра. Коли збудження нейрона співпадає з вдихом, то такий нейрон відноситься до інспіраторних, коли з видихом – до експіраторних. Нейрони цих ядер широко контактують з ретикулярною формацією мозкового стовбура.
І. Ядра бульбарного центру дихання:
Дорсальне ядро - має нейрони, які збуджуються при інспірації. Збудження нейронів дорсального ядра гальмує експіраторні нейрони вентрального.
А) І альфа –нейрони – збуджуються тільки під час вдиху (інспіраторні нейрони).
Імпульси від них передаються на мотонейрони діафрагми, розташовані в спинному мозку (3 і 4 шийні сегменти).
Б) І бета-нейрони – збуджуються одночасно з І альфа й при паузі останніх. Їх збудження гальмує активність І альфа-нейронів (запускають видих).
2. Вентральні ядра – їх верхня частина має більше інспіраторних нейронів, а нижня – експіраторних. Більша частина нервових волокон від цих ядер йде в грудні сегменти спинного мозку до мотонейронів міжреберних м’язів і м’язів живота (до м’язів вдиху або видиху), лише 20-25% волокон розгалужуються в ділянці діафрагмальних ядер.
ІІ. Пневмотаксичний центр – в передній частині варолієва моста. Він впливає на бульбарний центр, бере участь в регуляції зміни вдиху і видиху, в тривалості вдиху і видиху.
ІІІ. Апнейстичний центр – в нижніх відділах варолієва моста. Імпульси центру збуджують І бета нейрони дорсального ядра. При деяких хворобах виникає апнейстичне дихання: тривалі судомні вдихи, які перериваються короткими видихами.
ІІ. Рецептори судин і ЦНС в регуляції дихання:
Центральні хеморецептори – на вентральній поверхні довгастого мозку. Фактор подразнення цих рецепторів є підвищення концентрації Н+ у позаклітинній рідині, склад якої визначається метаболізмом оточуючих нейронів і місцевим кровотоком. Центральні хеморецептори стимулюють інспіраторні і експіраторні нейрони – посилюється вдих і видих. Тому при зниженні рН спинномозкової рідини лише на 0,01 вентиляція легень збільшується на 4 л/хв..
Периферичні хеморецептори – находяться в каротидних тільцях, які розташовані в ділянці біфуркації загальних сонних артерій і в аортальних тільцях, які розташовані на верхній і нижній поверхні дуги аорти.
А)Каротині тільця контролюють газовий склад крові, яка поступає до мозку. Чим нижчий Р О2 в крові, яка омиває рецептори, тим більше частота імпульсів, які йдуть від них до дихального центру. Імпульсація від каротидних хеморецепторів досягає інспіратор них нейронів довгастого мозку й затримує виключення вдиху, тобто поглиблює дихання. Збуджуються ці рецептори також при зниженні рН або при підвищенні напруги СО2 в артеріальній крові. Гіпоксія й гіперкапнія взаємно посилюють імпульсацію від цих рецепторів.
Б)Аортальні хеморецептори – менш регулюють дихання, більш відома їхня роль в регуляції кровообігу.
Рецептори легень і дихальних шліхів в регуляції дихання:
Механорецептори=рецептори розтягнення – в гладких м’язах повітроносних шляхів. Реагують на різну ступень розтягнення легень. Аферентні імпульси йдуть в дорсальне ядро бульбарного відділу дихального центру - активують І бета- нейрони. В свою чергу І бета -нейрони гальмують активність І альфа - нейрони, зупиняють вдих. Але така реакція виникає при високій частоті імпульсів. При низькій частоті імпульсів рецептори розтягнення навпаки подовжують вдих і вкорочують видих. Так, якщо штучно роздути легені, то вдих рефлекторно загальмується. Навпаки. зменшення об’єму легень призводить до поглиблення видиху. Цей рефлекс зветься рефлеком Герінга-Брейера. Імпульси цього рефлексу поступають із механорецепторів легень, потім імпульси йдуть по волокнам блукаючого нерва до бульбарного дихального центру. Він запобігає у дорослих перерозтягненню легень при вдиханні більше 1,5 л повітря (посилене дихання), у новонароджених приймає участь в забезпеченні періодики дихання в спокою.
Іритантні рецептори – в епітеліальному шарі повітроносних шляхів, особливо їх багато в коренях легенів. Їхні центри знаходяться в довгастому мозку. Іритантні рецептори володіють властивостями механорецепторів – збуджуються при значному зменшенні або збільшенні об’єму легень, і хеморецепторів – подразнюються парами їдких речовин (аміак, тютюн), холодним повітрям, БАВ (гістамін).
Подразнення іритантних рецепторів викликає:
захисні рефлексторнв реакції (кашель, чхання): скорочення видиху і видалення чужорідних частинок,
скорочення гладких м’язів бронхів: подразнення іритантних рецепторів, передача імпульсів через блукаючий нерв на монхів.Це спостерігається, наприклад, під час подразнення іритантних рецепторів гістаміном при бронхіальній астмі (бронхоспазм). Фізіологічне значення вказаного рефлексу – при вдиханні токсичних речовин зменшується бронхіальний просвіт та вентиляція легень і завдяки цьому токсичні речовини в меншей кількості потрапляють у кров.
Юкстамедулярні рецептори (j – рецептори) – розташовані в стінках альвеол
біля капілярів. Подразнюються вони при вступу БАВ в мале коло кровообігу, при збільшенні об’єму інтерстеціальної рідини легень. Імпульси від них йдуть в довгастий мозок по блукаючому нерву. У здорової людини ці рецептори знаходяться в стані легкого тонічного збудження. Посилення імпульсації призводить до частого поверхневого дихання. Вони разом з іритантними рецепторами викликають задишку при набряку легень.
Рецептори плеври – відносять до механорецепторів (рецептори розтягнення) й хеморецепторів. Вони відіграють роль при зміні характера дихання при порушенні гладкості листків плеври. Одночасно виникає відчуття болю, що пов’язано з рецепторами парієтального листка.
Рецептори носової порожнини – реагують на механічні і хімічні подразники. Вони подібні іритантним рецепторам (чхання, кашель, звуження бронів).
Рецептори дихальних м’язів – замикаються на рівні відповідних відділів спинного мозку і стовбуру. Фізіологічне значення – при утрудненні дихальних рухів автоматично посилюється сила скорочення м’язів.
Рецептори інших органів – активують ретикулярну формацію, а дихальний бульбарний центр є однією із структур РФ.
ІІІ. Дихання за різних умов.
Дихання при фізичному навантаженні. Потрібно підвищеної кількості кисню, вентиляція легень збільшується в 10-20 разів. До початку виконання роботи включаються умовно-рефлекторні механізми регуляції, які полягають в тому, що кора великих півкуль, викликаючи довільні рухи, активізує діяльність ДЦ. В організм надходить більше кисню і з’являється можливість почати інтенсивну м’язову діяльність. Крім цього, вентиляція легень посилюється рефлекторно під впливом імпульсів, які надходять від механорецепторів м’язів, що скорочуються. Під час фізичної роботи підвищується чутливість центральних хеморецепторів ДЦ і периферичних хеморецепторів каротидного синусу та дуги аорти до гіпоксії, гіперкапнії і зниження рН. При інтенсивному фізичному навантаженні особливо знижується рН внаслідок накопичення молочної кислоти, що посилено утворюється у працюючих м’язах.
Дихання при зниженому атмосферному тиску. При піди манні на висоту людина опиняється в умовах зниженого тиску, внаслідок цього в організм надходить менше кисню, розвивається гіпоксія. Вже на висоті 2,5-3 км над рівнем моря розвивається гіпоксія, яка стимулює каротині хеморецептори. При цьому настає гіпервентиляція – дихання глибоке і часте. При подальшому підніманні на 4-5 км відбувається зниження атмосферного тиску і розвивається висотна хвороба, причиною якої є різке зниження напруги СО2 в крові (виводиться при гіпервентиляції). Оскільки СО2 є подразником хеморецепторів, при його зниженні глибина дихання різко зменшується. Гіпоксія призводить до слабкості, підвищення АТ, головного болю, втраті свідомості. Дихання чистим киснем дозволяє людині зберегти працездатність навіть на висоті 11-12 км. При тривалому перебуванні в гірській місцевості розвивається акліматизація до гіпоксії. У таких людей збільшується кількість еритроцитів і Нв в крові, підвищується стійкість нервових клітин до гіпоксії, збільшується вентиляція легень.
Дихання при підвищеному атмосферному тиску – в цих умовах працюють водолази. При зануренні під воду збільшується кількість газів, розчинених у крові. Підвищення рівня О2 в крові може призвести до кисневого отруєння, яке супроводжується судомами. Тому при занурені на великі глибини застосовують гелієво-кисневі суміші. Особливої уваги потребує підйом водолазів від високого тиску на глибині до нормального на поверхні. При швидкому підйомі може розвинутись кесонова хвороба, зумовлена виходом азоту із тканин і закупоркою дрібних судин (газова емболія), що порушує циркуляцію крові і може призвести до смерті. Для лікування кесонної хвороби потерпілого поміщають в камери з високим тиском і цим викликають розчинення бульбашок азоту.
ІУ. Механізм першого вдиху новонародженого.
Газообмін плода відбувається через плаценту. Під час пологів плацентарний газообмін порушується. Це призводить до значного зниження О2 у крові дитини (гіпоксія) і збільшення СО2 (гіперкапнія). Вирішальним моментом є перев’язування пуповини при пологах. У цей час різко зростає гіперкапнія у крові новонародженого, що є сильним подразником хеморецепторів. Імпульсація досягає ДЦ і збуджує інспіраторні нейрони. Таким чином відбувається перший вдих. Інспіраторні нейрони також збуджуються під впливом нервових імпульсів, що надходять від механо- та терморецепторів шкіри.
Лекція №11.
Тема: «Травлення в ротовій порожнині і шлунку».
План:
І. Основні функції системи травлення.
ІІ. Ферменти. Типи травлення: порожнисте, мембранне, внутрішньоклітинне. Механізм регуляції травлення.
ІІІ. Слина: склад і властивості, механізм секреції та регуляції.
ІУ.Жування, ковтання. Фази. Регуляція.
У.Секреторна діяльність шлункових залоз.
Шлунковий сік. Механізм секреції та регуляція виділення. Фази регуляції секреції: мозкова, шлункова, кишкова.
УІ. Моторна функція шлунка. Її регуляція.
І. Функції ШКТ:
1)Моторна – відбувається за рахунок м’язової оболонки, забезпечення жування, ковтання, проштовхування їжі й виділення неперетравлених часточок їжі.
2)Секреторна – вироблення травних соків: слини, шлункового, підшлункового, кишкового і жовчі.
3)Інкреторна – утворення в травному каналі ряду гормонів, які впливають на процес травлення.
4)Екскреторна функція – виділення травними залозами в порожнину ШКТ продуктів обміну речовин, води, солей, ліків, потім їх виділення із організму.
5)Всмоктувальна функція – забезпечується слизовою оболонкою ШКТ.
6)Бактеріальна флора – вливає на імунітет, на обмін речовин. На синтез вітамінів.
ІІ. Ферменти. Типи травлення: порожнисте, мембранне, внутрішньоклітинне. Механізм регуляції травлення.
В травному каналі за допомогою ферментів і механічних процесів відбувається травлення їжі, тобто розщеплення складних полімерних сполук на складові частини – мономери. Ферменти травної системи – це біологічні каталізатори, які діють на кисневі зв’язки молекул білків, жирів і вуглеводів. Цей процес подрібнення великих молекул на дрібніші складові частини називають гідролітичним розщепленням, а ферменти – гідролазами. Залежно від дії ферменти травлення поділяються на протеолітичні (діють на пептидні зв’язки і розщеплюють білки), амілолітичні (діють на глюкоїдні зв’язки і розщеплюють вуглеводи), ліполітичні (діють на ефірні зв’язки і розщеплюють ліпіди).
Характерними особливостями ферментів є: специфічність – фермент діє на певну речовину; чутливість до реакції середовища; залежність від температури; більшість з них перебуває у неактивному стані й активуються або гідролітичними ферментами, або електролітами та окремими іонами (Са, цинк, магній).
Ферменти синтезуються залозистими клітинами. Гідроліз поживних речовин проходить у порожнинах шлунку та кишок, куди надходять травні соки. Особливо інтенсивна дія ферментів відбувається на цитолемах клітин слизової оболонки кишок, де вони розташовуються в певній послідовності. Цей феномен був названий в певній послідовності. Цей феномен був названий О.М.Уголевим пристінковим, або мембранним травленням. У слизовій оболонці ШКТ є клітини, які виділяють слиз мукопротеїдного походження, що захищає клітини травного каналу від само перетравлення ферментами, а також полегшує проходження їжі.
Типи травлення:
Внутрішньоклітинне – за рахунок потрапляння речовин ендоцитозом в ентероцити, і перетравлення речовин клітинними ферментами до кінцевих речовин.
Позаклітинне – порожнинне (дистанційне)=20-50% та мембранне (пристінкове) =50-80%. Порожнинне травлення особливо ефективне при розщепленні великих харчових молекул, цілих клітин ферментами.
Мембранне – на межі між внутрішньоклітинним і позаклітинним травленням, тобто на мембрані. На мембранах мікроворсинок відбувається розщеплення проміжних продуктів гідролізу (олігомери) до мономерів (кінцевих продуктів). Розщеплення у мікроворсинках відбувається тільки тоді, коли розміри молекул харчового субстрату менші розмірів пор оболонки мікроворсинок.
Особливості мембранного травлення: а) за рахунок ферментів, які мають подвійне походження: секретуються клітинами і абсорбуються вмістом кишки, б)ферменти, які фіксуються на клітинній мембрані таким чином, щоб активний центр був направлений в порожнину, в) відбуваються в стерильних умовах,
г) є заключним етапом в обробці їжі,
д) зближує процеси розщеплення і всмоктування за рахунок того, що кінцеві продукти переносяться на транспортних білках.
Регуляції процесів травлення.
Існують три типи ефекторних впливів регуляторних механізмів:
Функціональні – запуск, зміни, корекція тої чи іншої функціональної активності органу,
Судинорухомі – зміни рівня кровопостачання адекватній функціональній активності органу,
Трофічні – різноманітні зміни трофічних функцій в секреторних клітинах, приведення їх в співвідношення з конкретним рівням функціонального стану органу..
Види регуляції:
1. Нервово-рефлекторна – регуляція комплексом безумовних і умовних рефлексів. Безумовні рефлекси починаються з подразнення рецепторів ротової порожнини і органів ШКТ: а) місцевими рефлексами - рефлекторні дуги замикаються в самому органі або поряд, б) рефлексами за участю різних структур ЦНС, за допомогою «мозкового травного центру».
Центри травлення локалізовані в корі, в інших відділах. Особлива роль – стовбура мозку – там починається блукаючий нерв (сенсорний та ефекторний нерв ШКТ). Ефекторними шляхами рефлекторних впливів є симпатичні і парасимпатичні нерви. Умовно-рефлекторна регуляція формується в процесі розвитку організму на вигляд, запах їжі, час прийому, оточення. УР виражені найбільш в верхньої частині ШКТ. Так. УР в найбільшій мірі регулюють виділення слини, декілька менше – соку шлункових залоз, печінки, підшлункової залози.
2.Гуморальна регуляція – гормонами ендокринних залоз, власними гормонами ШКТ (гастроінтестінальними: гастрин, секретин). ГІГ розташовані дифузно в слизовій ШКТ, підшлункової залозі. Стимулюються вони під впливом хімічних і механічних подразників їжі. Гормони впливають на трофічні функції, на рухові, на всмоктування. В ШКТ утворюються також інші БАР: кініни, простагландіни (регулюють кровоток, моторику, секрецію).
ІІІ. У ротовій порожнині відбувається механічна обробка їжі (подрібнення їжі і змочування слиною), хімічна обробка і формування травної грудки:
Жування – подрібнення їжі зубами, жування здійснюється жувальними м’язами. Жування – рефлекторний акт. Їжа в роті подразнює рецептори, по аферентних волокнах трійчастого нерва подразнення передається в центр жування у довгастому мозку, а звідти по еферентних волокнах трійчастого нерва – до жувальних м’язів.
Слиноутворення і слиновиділення . Слина утворюється слинними залозами. Інтенсивність секреції – від 1,0 до 200,0 мл/год. В середньому за добу виділяється 500-2000мл слини. Склад слини: вода(99%), неорганічні речовини (солі кальцію, калію), органічні речовини (мукополісахариди, глікопротеїни, білки), муцин (ослизнює харчову грудку і сприяє ковтанню), ферменти (амілаза- розщеплює крохмаль до мальтози, мальтаза – розщеплює мальтозу до глюкози), лізоцим (бактерицидна речовина). Харчова грудка знаходиться в ротовій порожнині 15 с, тому дія ферментів слини продовжується в шлунку (20 хв).
Слина, що виділяється привушними залозами, серозна (мало муцину), під’язиковими – слизова, а підщелепними – змішана. Кількість і якість слини залежить від характеру їжі. Чим сухіша їжа, тим більш слини декретується.
Регуляція слиновиділення.
1)Умовно-рефлекторна – при дії умовних подразників (запах, розмова або думка про їжу). Рефлекторна дуга УР замикається в корі великих півкуль.
2)Безумовно-рефлекторна – при потраплянні їжі в рот. Збудження по аферентних волокнах трійчастого, лицевого, язикоглоткового та блукаючого нервів передається в парасимпатичний центр слиновиділення (в довгастому мозку). Звідси збудження по еферентних волокнах, що входять до складу тих самих нервів, направляється до слинних залоз. Симпатична іннервація слинних залоз здійснюється від бічних рогів ІІ-ІУ грудних сегментів спинного мозку. Подразнення парасимпатичних нервів призводить до утворення рідкої слини, а симпатичних нервів – густої слини.
3. Ковтання- рефлекторний акт. Ковтальний рефлекс починається з подразнення рецепторів кореня язика, піднебіння або задньої стінки глотки. До центру ковтання (в довгастому мозку) збудження поступає по язикоглотковому нерву. Еферентними шляхами є під’язиковий, язикоглотковий, трійчастий і блукаючий нерви, які йдуть до м’язів порожнини язику, глотки, гортані, стравоходу.
Фази ковтання:
Начальна – скорочення посмугованих м’язів (свавільне й несвавільне).
Глоткова – несвавільні (перистальтика). Швидка фаза.
Стравохідна - несвавільні (перистальтика). Повільна фаза (3-8 с).Скорочення циркулярних м’язів над стравоходом проштовхує її в нижню ділянку. Яка розширюється за рахунок скорочення повздовжніх м’язів.
Харчова грудка не потравляє в дихальні шляхі – цьому сприяє рефлекторне підняття м’якого піднебіння, що перекриває носоглотку, опускається надгортанник – перешкоджає вхід в гортань.
У.Секреторна діяльність шлункових залоз.
Шлункові залози поділяються на власні, кардіальні, пілоричні.
1)Власні залози шлунка – розташовані в ділянці тіла й дна. Вони побудовані із головних клітин (виробляють пепсиногени), парієнтальних = обкладкових (виробляють соляну кислоту), додаткові клітини (виробляють слиз). Їх сумарна діяльність забезпечує секрецію шлункового соку (2-2,5 л). Шлунковий сік – безколірна, прозора рідина, яка містить соляну кислоту. Його рН становить 1,5-1,8. Склад соку:
А) пепсиноген – профермент, який у порожнині шлунка перетворюється на активну форму ферменту – пепсин, що має здатність гідролізу вати (розщепляти) білкм;
Б) Хімозин – розщеплює білки молока (звурдження), виробляється переважно у дитячому віці;
В)Желатиназа – розщеплює сполучної тканини – желатин.
Г) НCl – викликає денатурацію і набубнявіння білків, сприяє їх розщепленню пепсинами, створює кисле середовище у шлунку( в ньому активні пепсини), звурджує молоко, регулює потрапляння харчової грудки в кишку, має бактерицидну дію, бере участь в регуляції вироблення слизової оболонки 12-палої кишки гормону секретину і ферменту ентерокінази. Шлунковий сік вміщує 0,4-0,5 % соляної кислоти (нормальна кислотність щлункового соку).
Д) Мукоїд або муцин (слиз) – утворює оболонку, яка захищає слизову від хімічних і механічних подразнень.
Є) Внутрішній фактор Касла – виробляється обкладовими клітинами. Він забезпечує всмоктування вітаміну В12. При відсутності фактора виникає анемія.
Ж) Ліпаза – гідроліз жирів молока.
2) Кардіальні і пілорічні залози розміщені в однойменних ділянках шлунка. Їх клітини продукують слиз та ферменти-дипептидази. До їх складу також входять значна кількість ендокринних клітин.
Регуляція виділення шлункового соку.
Нервові механізми
Гуморальні механізми
Розрізняють 3 фази секреції:
І. Мозкова фаза – складнорефлекторна, тобто шлунковий сік виділяється умовно- і безумовно рефлекторно:
А) умовні рефлекси – викликають виглядом, запахом, словом про їжу;
Б) безумовні рефлекси – при потраплянні їжі в рот. Збудження надходить у довгастий мозок, а звідти – по блукаючому нерву – до шлунка, стимулюючи виділення «апетитного» шлункового соку.
ІІ. Шлункова фаза – починається з моменту надходження харчової грудки в шлунок. Вона зумовлена нервовими і гуморальними механізмами.
1) Нервовий механізм. Харчовий хімус, заповнюючи шлунок, подразнює механорецептори слизової, імпульсація по аферентним волокнах блукаючого нерва передається в секреторний центр довгастого мозку і звідти по еферентних волокнах в складі того ж блукаючого нерва повертається до залоз шлунка, стимулюючи їх діяльність.
2) Гуморальний механізм. Гуморальними подразниками є гастрит, ацетилхолін.
А) Гастрит виробляється клітинами слизової шлунка і активується тільки в кислому середовищі. Із шлунка цей гормон всмоктується в кров і через неї діє на залозистий апарат шлунка, активуючи його. Вивільнення гастрину посилюється продуктами гідролізу білків, екстрактивними речовинами м’яса й овочів.
Б) Гістамін – утворюється в слизовій шлунка, стимулює шлункову секрецію. Його виділення посилюється в присутності ацетилхоліну і гастрину. Дія гістаміну опосередковується також через кров.
ІІІ. Кишкова фаза – починається з моменту надходження першої порції кислого хімусу в дванадцятипалу кишку. Впливи з кишки на шлункову секрецію здійснюється через:
1)Нервові механізми – подразнення йдуть з механо- і хеморецепторів кишки через секреторний блукаючий нерв. Рефлекторно посилюється шлункова секреція, активується перетравлення хімусу.
2) Гуморальні механізми – регулюються гормонами ШКТ:
А)Секретин, холецистокінін-панкреозимін – гальмують секрецію залоз шлунка, але стимулюють виділення пепсиногену. Вони виділяються слизовою кишки, коли надходить кислий вміст шлунка в 12-палу кишку.
Б) Продукти гідролізу жиру – підвищують виділення холецистокініну-панкреозиміну, що також гальмує шлункову секрецію в кишковій фазі.
В) Ентерогастрин, серотонін, соматостатин – гальмівний вплив на секрецію залоз шлунка.
Г) Продукти гідролізу білків – стимулюють шлункову секрецію.
Д) Біль, негативні емоції, посилена фізична робота – гальмують виділення шлункового соку через симпатичну нервову систему.
УІ. Моторна функція шлунка. Її регуляція.
Моторна функція шлунка полягає у скороченні його гладких м’язів. Ця функція забезпечує депонування в шлунку прийнятої їжі, перемішування її із шлунковим соком, пересування до виходу в кишечник і порційну евакуацію хімусу в 12-палу кишку. При скороченні м’язових волокон шлунка змінюється його об’єм і форма. При наповненні шлунка (резервуарна функція або депо їжі) не виникає підвищення тиску, його м’язи розтягуються. Крім того, під час вживання їжі відбувається додаткове розслаблення шлунка під дією блукаючого нерва, що називають об’ємною адаптацією. Харчовий хімус знаходится в шлунку 3-10 год. Найдовше затримується жирна їжа, а білкова затримується довше, ніж вуглеводна.
Регуляція моторної функції:
Нервова – моторику шлунка стимулює блукаючий нерв. Симпатичний нерв, навпаки, гальмує.
Гуморальна – гормони ШКТ (мотилін і гастрин) підсилюють моторику, а золецистокінін-панкреозимін, шлунково-інгібуючий пептид та бульбогастрон гальмують її. Продукти гідролізу жиру, що надходять у кров, також гальмують моторику шлунка.
Види скорочення шлунку:
1.Тонічні скорочення шлунка – підтримують постійний внутрішньопорожнинний тиск і відповідність об’єму шлунка до об’єму їжі. Без їжі шлунок знаходиться в спавшемся стані.
2. Перистальтичні хвилі скорочення стінок шлунка - починається через деякий час після прийому їжі Починаються прокатуватися хвилі скорочення на великій кривизні в ділянці входження в нього стравоходу, а потім розповсюджується уздовж стінок у напрямку по пілоричній частині. Скорочення м’язів розповсюджується зі швидкістю 10-40 см/с.
Виділяють хвилі трьох типів:
1 тип – хвилі низької амплітуди тривалістю 5-20 с.
2 тип – хвилі більш високої амплітуди, тривалістю 12-60 с. Ці хвилі утримують тонус шлунка і сприяють повільному перетравленню хімусу, який прилягає до стінок з шлунковим соком. Затухають в пілоричному відділі.
3 тип – для пілоричного відділу. Хвилі високої амплітуди, носять пропульсивний характер і сприяють евакуації хімусу в 12-палу кишку.
Робота пілоричного сфінктеру (пілоричний затульний рефлекс) - харчова кашка переходить зі шлунка в 12-палу кишку порціями (6-12 мл). Кислий хімус шлунка підходить до пілоруса і подразнює рецептори слизової. Збудження йде в ЦНС і звідти – до пілоричного сфінктера, який при цьому відкривається. Таким чином, порція кислого хімусу потрапляє в лужне середовище дванадцятипалої кишки. Перехід хімусу відбувається доти, поки реакція в 12-палій кишці не стане кислою. Кислота подразнює рецептори слизової кишки і сфінктер рефлекторно закривається. Він залишається закритим до того часу, поки реакція в кишці не стане знову лужною завдяки її нейтралізації жовчю та кишковим соком. Тоді сфінктер знову відкривається і пропускає чергову порцію хімусу з шлунка.
Лекція №18.
Тема: «Фізіологія контактних аналізаторів».
План:
І. Структурно-функціональна організація смакової і нюхових сенсорних систем. Види смаків, механізм сприйняття. Класифікація запахів, теорії сприйняття.
ІІ. С.-ф. організація шкірної та рухової сенсорних систем.
ІІІ. С.-ф. організація ноцицептивної (больової) та протибольової систем. Рівні оброблення інформації. Фізіологічна роль. Фізіологічні основи знеболювання.









Лекція
«Вища нервова діяльність. Сигнальні системи. Типи ВНД»
План:
Особливості функціонування ВНД. Функціональна асиметрія кори великих півкуль.
Фізіологічні основи формування мови. Сигнальні системи.
Мовні центри. Фонація і артикуляція.
Мислення. Свідомість.
Типи ВНД.
ОСОБЛИВОСТІ ПСИХІЧНОЇ ДІЯЛЬНОСТІ ЛЮДИНИ
Основні закономірності умовнорефлекторної діяльності вищих тварин і людини є спільними. Але мозок людини від мозку тварин принципово відрізняється у функціональному відношенні. І. П. Пав-ловим було висловлено ідею про специфіку мозку людини. Учений запропонував при оцінці функціональних особливостей мозку людини виходити з характеру тих сигналів, які специфічні тільки-для людини. Для тварин умовними сигналами, які ведуть до утворення умовних рефлексів, є безпосередні подразнення та їх сліди, тобто всі екстеро- та інтероцептивні подразнення, які надходять у кору великого мозку. Це те, що і ми вважаємо відчуттями, враженнями, уявленнями про навколишній світ. За І. П. Павловим,. це перша сигнальна система дійсності, яка є спільною для людини-і тварин. Але завдяки слову (мові) сформувалась друга, спеціально людська, сигнальна система дійсності. Вона розвинулась завдяки праці, суспільно-трудовій діяльності, яка сприяла все тіснішому спілкуванню первісних людей між собою і утворенню суспільства. При цьому з'явилась потреба в звуковій сигналізації — членороздільній мові, завдяки якій стали можливими узгодження трудових рухів і взаємної підтримки. «Слово зробило нас людьми»,— писав-І. П. Павлов.
ФУНКЦІОНАЛЬНА АСИМЕТРІЯ КОРИ ПІВКУЛЬ ВЕЛИКОГО МОЗКУ
Відомості про локалізацію й функцію окремих відділів головного мозку людини було отримано відносно недавно. Основою цих досліджень є анатомо-фізіологічні особливості: висхідні й низхідні провідні системи мозку переходять на протилежну половину тіла, а тому ліва півкуля великого мозку відповідає за соматосенсорну чутливість і рух правої половини тіла, а права навпаки.
Найвірогідніші дані про латералізацію функцій м о з -к у вчені отримали при обстеженні хворих на епілепсію, в яких з лікувальною метою повністю перерізували мозолисте тіло, яке складається з мільйонів волоконець, котрі сполучають обидві півкулі великого мозку. Дослідження свідчать про те, що аналітико-синтетичні процеси, які відбуваються в асоціативних зонах кори лівої і правої півкуль великого мозку, здійснюються по-різному. Особливо наочно це проявляється стосовно функцій, пов'язаних із мовою. У правші ізольована ліва півкуля великого мозку функціонує майже так само ефективно, як і весь мозок, здатна забез- печити всі види мови, а права півкуля не може забезпечити нї усну, ні письмову мову. Однак ці уявлення не є абсолютними. Доведено, що центральні механізми, котрі відповідають за письмо й читання, у правші дещо залежать від способу написання. При позначенні слова літерами цей центр локалізований у лівій півкулі великого мозку; при написанні ієрогліфів, коли використовується спосіб креслення складів, слів або фраз, центр письмової мови локалізується в правій півкулі.
Описані міжпівкульні функціональні відмінності проявляються і при опрацюванні інформації, яка надходить структурами першої сигнальної системи. Правша краще сприймає інформацію правим вухом, а у лівші така закономірність не простежується. У більшості людей бінокулярний зір здійснюється переважно правим оком (60—70 %). Таким чином, у багатьох людей асиметрія мозку проявляється в двох формах: право- і лівостороння. Приналежність до правші чи лівші спадково запрограмована. Переучування в дитинстві лівши, у якого природжено центр мови розташований праворуч, призводить до одночасного перенесення центра мови (в цілому або частково) в ліву півкулю великого мозку, а це може істотно вплинути на мовну функцію мозку, зокрема, зумовити заїкання.
Висловлюють припущення, що ліва півкуля обробляє інформацію послідовно, у міру її надходження, права ж працює одразу з кількома входами, одномоментно й паралельно. Певно, інтуїція і прозріння є функціями правої півкулі великого мозку. Сьогодні наші уявлення про функції головного мозку, його лівої та правої половини збігаються з уявленнями про можливість виділення мислячого і художнього типу людей. Основою логічного способу мислення є ліва півкуля великого мозку, а образного, інтуїтивного — права.
СЛОВО ЯК УМОВНИЙ ПОДРАЗНИК
Слово, будучи умовним подразником, характеризується такими властивостями: а) воно є сигналом конкретних сигналів (сигнал сигналів); б) здатне до узагальнення різних конкретних, чутливих сигналів першої сигнальної системи (наприклад, у слові «дерево» узагальнено ознаки всіх дерев, незалежно від їх конкретної різниці); в) слово може відволікати (абстрагувати) від конкретної, дійсності, на цьому грунті формується абстрактне, логічне мислення людини.
До умовних сигналів другої сигнальної системи слід зарахувати не тільки ті слова, які чуємо, бачимо написаними, але й нотні знаки, міміку та жести глухонімих, які теж виконують функції позначення перших конкретних сигналів дійсності. Потужним подразником людського мозку є так звана внутрішня мова (сам до себе). Слово як умовний подразник другої сигнальної системи діє на людину своїм змістом (семантикою), а не звуковою характеристикою (акустичною оболонкою). Про це свідчить ряд експериментів (наприклад, вироблений умовний рефлекс на слово «доріжка» може бути викликаний його синонімом «стежка»; умовний подразник — цифра 5 — може бути замінений цифрами 2 + 3 (або 10:2 тощо). У тварин слово теж може служити умовним сигналом, але діє воно своєю акустичною характеристикою, а не змістом.
Таким чином, людина сприймає безпосередньо навколишній світ на основі першої сигнальної системи, а абстрактне, понятійне мислення зв'язане з другою сигнальною системою. Друга сигнальна система є вищою формою нервової діяльності людини, вищим регулятором людської поведінки.
Дуже важливо, що слово є не тільки фізіологічним, але й сильним лікувальним чинником. Тому лікарі та інші медичні працівники повинні бути дуже обережними в словах, щоб не спричинити у хворого так звану ятрогенію, тобто захворювання (переважно психоневроз), які виникають при недоречних висловлюваннях лікаря, іноді навіть відповідній міміці.
Фізіологічні основи мови. Мова є дуже складною психічною діяльністю, специфічно людською формою відображення дійсності.
Функції мови. Мова виконує важливі функції, а саме: комунікативну, понятійну, регуляторну й програмуючу. Комунікативну функцію можна визначити як процес спілкування за допомогою мови (в елементарній формі ця функція притаманна й тваринам у вигляді відтворення звукових або зорових подразників, які є сигналами для інших тварин). Понятійна функція полягає в тому, що мова є основою і засобом понятійного, абстрактного, логічного мислення, механізмом інтелектуальної діяльності людини. Регуляторна функція мови полягає в тому, що за допомогою словесних інструкцій або внутрішньої мови здійснюється керування найрізноманітнішими власними мимовільними діями і рухами, а також поведінкою інших людей. Слова впливають також на діяльність різних органів і систем організму людини. Програмуюча функція мови виражається в побудові (за допомогою внутрішньої мови) змістовних схем висловлювання, граматичної структури речення, різних рухів і дій.
Види мови. Виділяють експресивну і імпресивну мову. Експресивна мова — це процес мовного висловлювання (у вигляді усної мови чи письма), який починається із задуму (програми висловлювання), потім проходить стадію внутрішньої мови і завершується висловлюванням за допомогою слів. Імпресивна мова — процес розуміння мовного висловлювання (усного чи письмового), який розпочинається із сенсорного його сприйняття (через слух або зір), потім відбувається декодування словесної інформації і завершується формуванням змістової її схеми. Для декодування словесної інформації потрібні активний аналіз найістотніших елементів її змісту, вилучення їх із пам'яті. Ці процеси здійснюються переважно скроне-тім'яно-потиличними відділами лівої півкулі великого мозку. Ушкодження цих відділів спричинюють розлади розуміння загального змісту мови, складних логіко-граматичних форм мови і лічильних операцій.
Усі ці форми мови складають єдину функціональну систему. Про це свідчать як дані нейропсихології, так і клінічні спостереження: при локальних ушкодженнях великого мозку (переважно лівої півкулі) розлади цоширюються на всі форми мовної діяльності з переважанням того чи того дефекту мови. Такі мовні розлади називають афазіями.
Центри мови займають кілька ділянок кори великого мозку. Понад 100 років тому П. Брока (1861) вперше виявив, що при ураженні нижніх відділів третьої лобної лівої півкулі втрачається мова. Цей центр локалізується поруч із рухливою ділянкою, котра контролює функцію м'язів язика і глотки, обличчя, тобто тих м'язів, які беруть участь у артикуляції. Він називається моторним центром мови. Екстирпація або інші ушкодження зони Брока призводять до афазії. Такі хворі розуміють мову, але говорити не можуть.
Другим центром мови є ділянка задніх відділів першої скроневої борозни (сенсорний центр мови Верніке). Він розташований поруч із слуховою зоною (мал. 246). Ураження його супроводжується, порушенням розуміння мови при збереженні здатності до перекрученої спонтанної мови. Цей тип патології називається сенсорною афазією.
Пізніше виникли центри мови, які забезпечують процеси читання й письма. Центр письмової мови розташований у ділянці потилиці — перед кірковим відділом зорового аналізатора. Однією із форм афазії є амнезія (потилична афазія), яка характеризується забуванням окремих слів (мовна пам'ять).
Перераховані центри мови в більшості людей містяться в лівій півкулі великого мозку. Локалізація центра мови в лівій півкулі великого мозку свідчить про те, що виникнення мови зв'язане з трудовою діяльністю людини: її досконаліша (вміла) права рука іннервується саме із лівої половини великого мозку а саме передньою лімбічною корою, покришкою і центральною сірою речовиною середнього мозку. Процеси, які забезпечують емоційне забарвлення і розуміння слова, є природженими.
УТВОРЕННЯ ГОЛОСУ (ВОКАЛІЗАЦІЯ)
Будова верхніх дихальних шляхів забезпечує не тільки проведення повітря в легені і назад, але й виконання однієї із найважливіших для людини функцій — утворення голосу. Це забезпечується: а) особливостями будови гортані; б) наявністю резонаторів; в) здатністю керувати швидкістю руху повітря; г) здатністю контролювати положення і натяг голосових зв'язок положенням губ, м'якого піднебіння.
Вокалізація включає два взаємопов'язаних процеси: фонацію і артикуляцію.
Фонація. Звук утворюється при вібрації голосових зв'язок тоді, коли повітря під час видиху проходить між ними. Голосові зв'язки розташовані вздовж латеральних стінок гортані. їх положення і натяг забезпечуються групою відповідних м'язів (мал. 247). Так, при скороченні заднього персневидно-черпакуватого м'яза, що рухає назад черпакуваті хрящі, голосові зв'язки натягуються. Поперечний черпакуватий м'яз при скороченні зближує голосові зв'язки. Латеральні персневидно-черпакуваті м'язи, зближуючи черпакуваті хрящі, розширюють голосові зв'язки.
Особлива роль належить щитовидно-черпакуватим м'язам, які містяться в складках разом із голосовими зв'язками. Ці м'язи складаються з ізольованих волокон, кожне з яких іннервується окремою гілочкою відповідного нерва. Завдяки цьому вони можуть скорочуватися окремими частинами і регулювати тонші голосові зв'язки й щілину, яка створюється між ними.
Вібрація голосових зв'язок відбувається при відносно швидкому про-• ходженні повітря під час видиху через звужену голосову щілину. Вважають, що вібрація відбувається завдяки такому механізму. При звужених голосових зв'язках повітря, яке проходить, їх роздимає. При цьому відносна вузькість щілини сприяє току повітря.в цьому відділі, що в свою чергу створює часткове розрідження в просторі між голосовими зв'язками. Завдяки цьому, згідно з законом Бернуллі, голосові зв'язки перекривають одна одну. Внаслідок цього потік повітря на деякий час припиняється, але під його тиском, який при цьому підвищується, вони знову розсуваються. Зазначений ефект повторюється, що і зумовлює вібрацію голосових зв'язок та виникнення звука.
Частота вібрації змінюється за рахунок послаблення або збільшення напруги голосових зв'язок та зміни конфігурації голосової щілини. Перший механізм забезпечується скороченням власне м'язів гортані. М'язи, які оточують гортань, теж можуть брати участь у зміні положення хрящів. При збільшенні напруги голосових зв'язок формується високий звук високого тону, а при їх зниженні створюються умови для формування низького тону. Висота звука залежить також від кута між зв'язками (мал. 248). Гострий кут забезпечує високочастотний звук, а при його збільшенні тон знижується.
Артикуляція. Основними органами артикуляції є губи, язик і м'яке піднебіння. їх положення під час проходження повітря дозволяє відтворювати окремі звуки. Стан кожного із них, конфігурація контролюються свідомо корою великих півкуль.
Індивідуальні особливості губ, язика, м'якого піднебіння надають звукам індивідуального забарвлення. Велике значення при цьому мають резонатори — порожнини рота, носа, пазухи лицьового черепа, глотки та грудної клітки.
Міжцентральні зв'язки в забезпеченні формування мови. У процесі формування мови виникає взаємодія між окремими її центрами. Вона здійснюється за двома зв'язками: прямим кортико-кортикальним і опосередкованим через таламус. Різні структури кооперуються за такою схемою. Спочатку акустична інформація, котра міститься в слові, обробляється в слуховій сенсорній системі мозку. Потім вона передається у розташований поруч із кірковим краєм слухового аналізатора центр Верніке, де розшифровується зміст слова. При прочитанні слова імпульси, які надходять у зорову сенсорну систему, заносяться в кутову борозну, через яку зорове нейронне збудження сполучається з його акустичним аналогом, розташованим у зоні Верніке. Для вимовлення слова треба, щоб уявлення про його зображення надійшло в зону Брока. -Зв'язуються названі центри за допомогою дугоподібного жмута волокон. У ділянці Брока імпульси, які надходять, сприяють формуванню програми артикуляції. Ця програма реалізується завдяки тіередачі імпульсів у обидві половини моторної зони мозку (пре-центральну борозну). Але для вимовлення звуків потрібне узгодження рухів відповідних органів з проходженням повітря.
Окрім цього, мовна функція має ще одну особливість — її емоційне забарвлення. Афективність мови забезпечують зв'язки лобної і тім'яно-скроневої часток із структурами лімбічної системи,
МИСЛЕННЯ
Мислення є вищою формою відображення світу. Це процес опосередкованого, узагальненого відображення дійсності; активна психічна діяльність, спрямована на розв'язання певної задачі (досягнення мети). Мислення виникає за наявності відповідного мотиву. Як зазначалось вище, розумова діяльність значною мірою опосередковується мовною функцією (внутрішня мова). Згідно із даними нейрофізіологічних та нейропсихологічних досліджень, наочно-образне або предметно-просторове мислення обумовлюється переважно структурами правої півкулі великого мозку, а абстрактне, словесно-логічне — лівої. Порушення мислення, інтелектуальної діяльності спостерігається загалом при ушкодженні тім'яно-потиличних та скроневих відділів лівої півкулі великого мозку лобних часток.
Свідомість — специфічна форма відображення діяльності й цілеспрямованого регулювання взаємовідносин особи з навколишнім світом. Вона формується тільки за умов суспільного життя. Свідомість визначається також як знання, яке за допомогою слів, математичних символів і узагальнюючих образів художніх творів може бути передане іншим людям, стати надбанням інших членів суспільства. Вона об'єднує все те, що може бути повідомлено або повідомляється, передається іншим людям. У формуванні свідомості вирішальну роль відіграє діяльність мовних структур мозку. Як зазначалося вище, для усвідомлення потрібен деякий рівень ться висхідною активізуючою системою головного мозку (ретикулярна формація мозкового стовбура, неспецифічні ядра таламуса, гіпоталамо-лімбічні структури).
Поряд із свідомістю психічна діяльність людини включає в себе підсвідомість. Термін «підсвідомість» (замість «несвідоме») використовується для позначення неусвідомлюваного психічного, нерозривно пов'язаного із свідомістю. До цієї групи психічних явищ належить усе те, що було усвідомленим або може стати усвідомленим за певних умов, наприклад, глибоко й міцно засвоєні людиною соціальні норми («голос совісті», «почуття обов'язку», «поклик серця» тощо), міцно автоматизовані навики (наприклад, рухові навики шофера, спортсмена, піаніста та ін.), поява інтуїції, яка грунтується лише на раніше набутому життєвому або професійному досвіді. Так, досвідчений лікар інтуїтивно (підсвідомо) ставить правильний діагноз, тому що його мозок зберігає сліди (є н г р а м и) ряду симптомів, типових для даного захворювання (характерна поза, міміка, колір шкіри, забарвлення склер тощо). Специфічними проявами діяльності підсвідомості є активізація творчих можливостей людини у стані глибокого гіпнозу, гіпнопедія (навчання під час сну, тобто без участі свідомості), а також, очевидно, і механізм сновидінь як різновидності пошукової активності. Про підсвідомі прояви психічної діяльності людини свідчить також можливість вироблення умовних рефлексів у відповідь на невідчутні (підпорогові) звукові подразнення.
Різновидом несвідомого психічного є також надсвідомість (су-персвідомість). До сфери надсвідомого належать первинні етапи будь-якої творчості (виникнення нових ідей, сміливих гіпотез, оригінального задуму, творчих задумів),.які виникають на грунті слідів, отриманих іззовні вражень (без свідомого вольового зусилля, передчасного втручання свідомості тощо). Надсвідомість відіграє важливу роль і у походженні різноманітних забобонів, міфів та ін.
Патологічно змінена взаємодія свідомості з під- і надсвідо-містю може проявитися у вигляді марення, галюцинацій, які виникають під впливом деяких психотропних речовин (ЛСД, гало-перидол та ін.).
Таким чином, психічна діяльність людини має трьохрівневу структуру, включаючи в себе свідомість, підсвідомість і надсвідомість.
ІНДИВІДУАЛЬНІ [ТИПОЛОГІЧНІ) ОСОБЛИВОСТІ ЛЮДИНИ
При аналізі індивідуальних особливостей людини, які проявляються в її соціальній поведінці, звичайно користуються такими термінами, як «темперамент», «характер» та «особистість».
Темперамент (у пер. з грецьк.— пропорція), або тип нервової системи, характеризує природжені властивості і особливості психічної діяльності індивіда (генотип). У структурі темпераменту можна виділити три головних компоненти — загальну активність індивіда, його рухові прояви й емоційність. Характер — індивідуальний склад психічного життя, форма його поведінки, «норов» людини, який проявляється в її манерах, звичках, емоційній сфері. Характер є основою поведінки людини. Формується він під впливом конкретного соціального середовища, умов виховання. Особистість може бути визначена як окремо взята конкретна людина. Особистість включає в себе і темперамент, і характер, і інтелект людини, а також соціально значущі риси, які характеризують індивіда як члена того чи того суспільства або спільності людей. Особистість інтегрує різні психічні процеси і формується (як і характер) під впливом конкретного соціального середовища. Поняття «характер» і «темперамент» часто ототожнюються, хоча характер є не стільки вираженням біологічних, природних властивостей нервової системи, скільки наслідком впливу на людину навколишнього середовища, насамперед суспільних, соціальних умов.
Класифікація темпераментів.
Ще Гіппократом була створена класифікація темпераментів, у основу якої було покладено сукупність постійних ознак у поведінці людини, його життєву активність, ініціативу й здатність до більш-менш енергійної діяльності. Великий вчений вважав, що надлишкове виділення крові зумовлює темперамент сангвінічний, надмірна продукція жовтої жовчі — холеричний, чорної жовчі — меланхолічний, слизу — флегматичний. Це уявлення про механізм виникнення темпераментів у світлі сучасних знань є наївними, але вчення про темпераменти і їх систематизація не втратили свого значення й досі.
Вивчаючи закономірності діяльності великого мозку, І. П. Павлов переконався у можливості оцінювати індивідуальні особливості його функцій на підставі вчення про типи нервової системи. В основу своєї класифікації типів нервової системи, або темпераментів, І.П. Павлов поклав такі головні принципи: силу, врівноваженість і рухомість нервових процесів (збудження і гальмування) у корі великого мозку.
Під силою збуджувального процесу розуміють здатність переносити досить сильні дії без вмикання позамежного гальмування. Сила гальмівного процесу характеризується швидкістю і міцністю внутрішнього (диференціювальне, запізніле тощо) гальмування. Сила нервових процесів у одного й того ж індивіда не є постійною, вона залежить від віку, харчування, ендокринних впливів, тренування нервової системи тощо.
Ступінь урівноваження процесів збудження та гальмування. Якщо вони врівноважують один одного і навіть при сильному нервовому напруженні цей баланс нервових процесів зберігається, то такий тип нервової системи називається врівноваженим. Якщо ж один із процесів домінує (частіше переважає процес збудження над процесом гальмування), то такий тип називається неврівноваженим.
Рухливість нервових процесів- здатність швидко або повільно, легко або важко збуджуватись чи гальмуватись.
Таким чином, оцінюючи такі вказані вище фізіологічні властивості нервової системи, І. П. Павлов виділів чотири основні типи нервової системи вищих тварин і людини:
1) сильний, урівноважений, рухомий («жвавий»);
2) сильний, урівноважений, інертний («спокійний», «солідний»);
3) сильний, неурівноважений з переважанням процесів збудження над процесами гальмування і з рухомою нервовою системою (нестримний);
4) слабкий (вирізняється слабкістю як збудливого, так і гальмівного процесів і схильністю до розвитку позамежного гальмування).
Класифікація темпераментів, запропонована Гіппократом, найближча до класифікації І. П. Павлова. При оцінці варіантів сильного типу треба враховувати, що сильний урівноважений тип з рухомою нервовою системою відповідає сангвінічному темпераменту, сильний урівноважений тип з інертною нервовою системою — флегматичному, сильний неврівноважений, нестримний тип — холеричному, слабкий тип — меланхолічному темпераменту.
Сангвінік вирізняється жвавістю, енергійністю, працездатністю, життєрадісністю, швидкою зміною станів, на зовнішні впливи реагує хоч і швидко, але неглибоко, добре пристосовується до життєвих умов (стереотипів). Холерик ще рухливіший від сангвініка, дуже енергійний (не знає міри!), але його життєва енергія швидко вичерпується і почата справа рідко доводиться до кінця. Він легко переходить від стану сильного збудження до млявості, часто вступає в конфлікти з людьми. Флегматики працездатні, повільні, холоднокровні. У них настільки слабка активність, що іноді межує з байдужістю. Для меланхоліка характерні загальна слабкість життєдіяльності, невпевненість у собі, нерішучість, відсутність ініціативи й активності, постійне незадоволення. Працездатність залежить від конкретних умов.
Характеризуючи форму поведінки людини, індивідуальний склад його духовного життя, ми часто говоримо про характер людини: сильний, слабкий, твердий, м'який, тяжкий, наполегливий тощо. Характер проявляється в діяльності людини (активний, цілеспрямований, нерішучий та ін.).
При формуванні типів нервової системи виняткового значення набувають взаємодія між першою і другою сигнальними системами та рівновага між ними. Цей принцип був покладений І. П. Павловим в основу класифікації спеціальних типів людини. Було виділено три основних типи — художній, мислячий і середній. Людина, що належить до художнього типу, добре сприймає навколишню дійсність (барви, звуки, запахи тощо, тобто конкретні сигнали). Такі люди охоплюють дійсність у цілому, повністю. Людина мислячого типу, навпаки, кожне явище аналізує, розбирає до найменших деталей, створює абстрактні уявлення про явища й події, які відбуваються. У людини, яка належить до художнього типу, особливо яскраве виявлена діяльність першої сигнальної системи (хоча у неї домінує вплив другої сигнальної системи). Для людини, яка належить до мислячого типу, характерні чітко виражені впливи другої сигнальної системи. У середнього типу певною мірою діють обидві сигнальні системи. Спеціально людські типи можна розглядати як наслідок індивідуальних варіацій функціональної асиметрії великого мозку. Художньому типові відповідає відносне домінування правої (немовної, конкретнообразної) півкулі, а мислячому — лівої.
У міру подальших досліджень типології людини ставало все очевиднішим, що перелік властивостей нервової системи, комбінація яких характеризує ту чи ту індивідуальність, слід істотно розширити. За допомогою спеціально розроблених психологічних тестів Г. Айзенк виділив три основні параметри: 1) екстра- та інтроверсивність; 2) емоційна стійкість і невротизм; 3) психотизм, протилежним полюсом якого є стійке дотримування соціальних норм. Екстраверт характеризується як відкритий, балакучий, соціальний, активний суб'єкт, а інтроверт — як замкнутий, пасивний, мовчазний. Невроїдний суб'єкт тривожний, схильний до гніву, емоційно нестійкий, йому протистоїть емоційно стійка особистість. Високопсихоїдний тип — егоцентричний, холодний, байдужий до всіх і агресивний суб'єкт. Ступінь екстра- та інтроверсивності визначається головним чином за допомогою спеціально розроблених анкет.
Вважають, що в основі екстраверсії лежать індивідуальні особливості взаємодії активізуючої ретикулярної формації, передніх відділів нової кори, У інтроверта більш розвинена септогіпокампальна, гальмівна система. Ступінь невротизму залежить від індивідуальних особливостей взаємодії лімбічних структур і нової кори.
Лекція № 9.
Тема: Нервова регуляція вегетативних функцій.
План:
Структурно-функціональна організація автономної нервової системи.
Симпатичний, парасимпатичний та метасимпатичний відділи. Їхня роль в регуляції
вісцеральних функцій. Автономні ганглії. Функції.
2. Механізм передачі збудження в синапсах симпатичної та парасимпатичної нервових
систем.
Вплив симпатичної, парасимпатичної та метасимпатичного відділів на функції організму.
Центральне регулювання в регуляції вісцеральних функцій.
Роль стовбура мозку та гіпоталамуса в регуляції вісцеральних функцій.