

14.2. Общая характеристика системы пищеварения
14.2.1. ПИЩЕВАРИТЕЛЬНЫЕ ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
А. Моторная (двигательная) функция — строго координированная сократительная деятельность исчерченных и гладких мышц пищеварительного тракта, обеспечивающая из-
мельчение пищи, ее перемешивание с пищеварительными секретами и перемещение пищевого содержимого в дистальном направлении. Моторика определяет продолжительность задержки пищевого содержимого в каждом отделе пищеварительного тракта, оптимальную для его механической и химической обработки и образования необходимого количества питательных веществ, их транспорта в кровь и лимфу, а также скорость перехода (эвакуации) частично переваренных пищевых веществ в дистально расположенный участок пищеварительного канала для последующей физической и химической обработки.
Строго координированная сократительная активность глад ко мы щечных элементов желчного пузыря, желчного и панкреатического протоков и их сфинктеров способствует выведению секретов печени и поджелудочной железы в просвет двенадцатиперстной кишки, где они ощелачивают кислый желудочный химус. Моторная деятельность тонкой кишки обеспечивает перемешивание и продвижение кишечного химуса по кишке, смену слоя химуса у ее слизистой оболочки, переход химуса из тонкой в толстую кишку. Моторика толстой кишки обеспечивает накопление кишечного содержимого, всасывание воды, формирование каловых масс и их удаление во внешнюю среду.
Б. Секреторная функция — совокупность процессов, обеспечивающих синтез секреторной клеткой специфического продукта — секрета из веществ, поступающих в клетку, и выделение его из клетки. Периодически повторяющиеся в определенной последовательности процессы, характеризующие деятельность секреторной клетки, носят название секреторного цикла, в котором различают три фазы: поглощение материала, внутриклеточный синтез и секретовыделение.
В составе секретов пищеварительных желез в полость пищеварительного тракта поступают ферменты, осуществляющие гидролиз пищевых веществ; электролиты, создающие оптимальный для гидролиза уровень рН; соли желчных кислот, обеспечивающие эмульгирование жиров и липидов; мукоид-ные вещества, выполняющие защитную роль.
По строению и характеру выделяемого секрета клетки пищеварительных желез подразделяют на три вида: белок-, мукоид- и ми-нералсекретирующие. Состав и свойства пищеварительного сока определяются количественными соотношениями активированных гландулоцитов разных видов. Количественные и качественные изменения секреции пи-
338
щеварительных желез адаптируются к виду принятой пищи.
В. Всасывательная функция заключается в переносе продуктов гидролиза, воды, солей и витаминов из полости пищеварительного тракта через слизистую оболочку во внутреннюю среду организма с помощью различных механизмов транспорта.
Ввиду кратковременности ферментативной обработки углеводов в ротовой полости, неглубокого гидролитического расщепления белков в желудке и отсутствия условий расщепления жиров в желудке всасывание в этих отделах пищеварительного тракта слабо выражено. Напротив, в тонкой кишке, где образуется основная масса конечных продуктов гидролиза белков, жиров и углеводов, интенсивность всасывания максимальна. Всасывание воды продолжается в толстой кишке, что необходимо для образования кала. Таким образом, все необходимые для жизнедеятельности организма вещества переходят через стенку кишечника в кровь и лимфу, а неусвоенные (неперевариваемые) компоненты пищи вместе с экскретами в виде экскрементов выводятся наружу.
14.2.2. НЕПИЩЕВАРИТЕЛЪНЫЕ ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
А. Защитная функция заключается в участии пищеварительного тракта в обеспечении неспецифических и специфических механизмов зашиты организма от чужеродных агентов и осуществляется с помощью нескольких механизмов.
-
Барьерные неспецифические механизмы зашиты обусловлены способностью слизис тых оболочек пищеварительного тракта пре пятствовать проникновению во внутреннюю среду организма пищевых веществ и патоген ных бактерий.
-
К неспецифическим факторам защиты относят также бактерицидное и бактериоста- тическое действие пищеварительных соков (Р.В.Петров).
-
Неспецифические клеточные механизмы защиты пищеварительного тракта определя ются фагоцитарной активностью лейкоцитов.
-
Неспецифические гуморальные механизмы защиты обеспечиваются способностью пи щеварительного тракта вырабатывать «есте ственные» антитела при контакте с облигат- ной кишечной микрофлорой.
-
Специфические клеточные и гуморальные механизмы защиты обеспечиваются иммуно- компетентными Т- и В-лимфоцитами мест-
ной иммунной системы пищеварительного тракта. Последняя включает миндалины глоточного кольца, солитарные лимфатические фолликулы в стенке кишки, пейеровы бляшки, червеобразный отросток, плазматические клетки слизистой оболочки желудка и кишечника.
Б. Метаболическая функция заключается в кругообороте эндогенных веществ между кровью и пищеварительным трактом, обеспечивающим возможность их повторного использования в процессах обмена веществ или пищеварительной деятельности.
В условиях физиологического голода эндогенные белки периодически выделяются из крови в полость желудочно-кишечного тракта в составе пищеварительных соков, где они подвергаются гидролизу, а образующиеся при этом аминокислоты всасываются в кровь и включаются в метаболизм. Такой кругооборот играет важную роль в механизме поддержания постоянства аминокислотного состава смеси, всасываемой в кишечнике, вне зависимости от колебаний аминокислотного состава пищевых белков (до тех пор, пока в организме имеются запасы незаменимых материалов).
Значительное количество воды и растворенных в ней неорганических солей циркулирует между кровью и пищеварительным трактом. Пищеварительные железы выводят из крови в ходе секреции электролиты, которые депонируются в содержимом желудочно-кишечного тракта и по мере необходимости вновь поступают в кровь и используются в метаболических процессах.
В. Экскреторная (выделительная) функция заключается в выведении из крови с секретами желез в полость пищеварительного тракта продуктов обмена и различных чужеродных веществ, поступивших в кровоток энтераль-ным или парентеральным путем. В пищеварительный канал экскретируются небелковые азотсодержащие вещества (мочевина, аммиак, креатинин, креатин), изотопы и красители, вводимые в организм с диагностическими целями, соли тяжелых металлов, лекарственные вещества. Доказана возможность элиминации слизистыми оболочками желудочно-кишечного тракта некоторых изотопов и красителей не только с жидкой частью пищеварительных соков, но и слизью. Роль экскреторных процессов в пищеварительном тракте особенно возрастает при недостаточной функции почек.
Г. Эндокринная функция заключается в секреции гормонов поджелудочной железой и специфическими клетками диффузной эн-
339
докринной системы пищеварительного тракта пептидов и аминов — гастроинтестиналь-ных гормонов, которые через кровь или местно (паракринным путем) оказывают регулирующие влияния на пищеварительные функции или другие системы организма.
Д. Непищеварительные функции печени см. раздел 14.6.3.
14.2.3. ОСОБЕННОСТИ РЕГУЛЯЦИИ ФУНКЦИЙ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
А. Факторы, запускающие деятельность пищеварительной системы. Натощак пищеварительный тракт находится в состоянии относительного покоя, для которого характерна периодическая функциональная активность. Прием пищи оказывает мощное пусковое влияние на проксимальные отделы пищеварительного тракта, что проявляется в виде интенсивной, но непродолжительной секреции слюнных, желудочных и поджелудочной желез, увеличения желчевыделения, пищевой релаксации (расслабление) желудка, кратковременного снижения моторной активности начальных участков тонкой кишки. Такие изменения секреции и моторики характерны для рефлекторных воздействий на систему пищеварения, причем пищеварительные секреты характеризуются высоким содержанием ферментов, накопленных в железах в периоды относительного покоя («запальный сок» по И.П.Павлову).
В дальнейшем интенсивность и характер секреторной и моторной деятельности органов пищеварительной системы изменяются за счет корригирующих нервных и гуморальных влияний по принципу обратной связи, формирующейся на основе рецепции содержимого пищеварительного тракта: его объема, консистенции, величины внутриполост-ного давления, наличия недостаточно измельченных кусочков пищи, рН, концентрации промежуточных и конечных продуктов гидролиза пищевых веществ, пищеварительных ферментов, температуры и осмотического давления.
Перечисленные факторы с помощью механизмов обратной связи (афферентная им-пульсация в ЦНС и в пределах интраорган-ной нервной системы, гуморальные вещества, действующие местно и через кровь) обеспечивают регулирующие влияния на органы пищеварительной системы с помощью ЦНС, интраорганной нервной системы и эндокринного аппарата самого пищеварительного тракта. Гуморальными афферентными сигна-
лами являются продукты гидролиза пищевых веществ (глюкоза, аминокислоты), экстрактивные вещества (например, капустный сок), пептид-гормоны диффузной эндокринной системы желудочно-кишечного тракта. Эти гормоны действуют на секреторные клетки непосредственно, а также с помощью ЦНС и интраорганной нервной системы. Эндокринные клетки желудка и кишечника имеют рецептивный полюс, обращенный в полость желудочно-кишечного тракта. Он взаимодействует со специфическими химическими веществами пищевого содержимого, что приводит к высвобождению гастроинтес-тинальных гормонов.
Наличие дублирующих каналов нервной и гуморальной обратной связи дает возможность пищевому центру получать достоверную и надежную информацию о многосторонней деятельности различных отделов пищеварительного тракта и достижении в процессе пищеварения этапных и конечного приспособительных результатов.
Нервная регуляция секреции и моторики осуществляется с помощью центральных, периферических и местных рефлексов (рис. 14.1-14.3).
Б. Вклад нервного и гуморального механизмов в регуляцию функций пищеварительной системы.
Нервный механизм. Влияние ЦНС наиболее выражено в начальном отделе пищеварительного тракта. Ее роль в регуляции секреции и моторики постепенно снижается вдис-тальном направлении по ходу пищеварительного тракта. Напротив, регуляторная роль интраорганной нервной системы в каудаль-ном направлении возрастает. В интраорганной нервной системе содержится значительное количество пептидергических нейронов, способных синтезировать и освобождать ВИП, соматостатин, нейротензин, вешество П, энкефалин, гастриносвобождающий пептид, холецистокинин. В интрамуральных ганглиях имеются как возбуждающие, так и тормозные нейроны. Основными возбуждающими нейронами являются холинергические. Торможение может быть связано с действием постганглионарных симпатических волокон на холинергические нейроны и непосредственно на эффекторные клетки. Другим видом торможения является неадренергическое не-холинергическое торможение, связанное с выделением пептидергическими нейронами тормозных медиаторов: ВИП, АТФ.
Гуморальные механизмы регуляции с наибольшей отчетливостью выражены в «средней» части пищеварительной системы (гас-
340
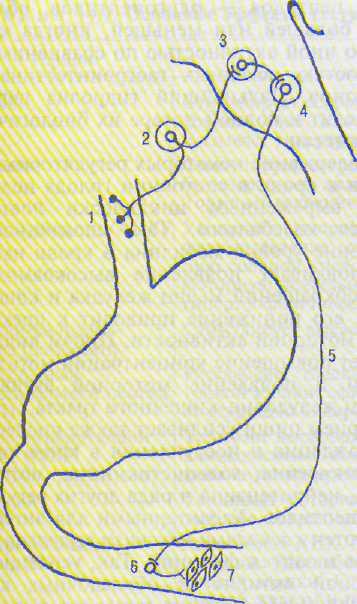
Рис. 14.1. Рефлекторная дуга центрального пище-водно-кишечного моторного рефлекса.
\ - механореиепторы пишевода; 2 — афферентный нейрон узловатого ганглия; 3 — чувствительное ядро продолговатого мозга; 4 — ядро блуждающего нерва (тела пре~ ганглионарных нейронов); 5 — преганглионарное парасимпатическое волокно: 6 — ганглионарный парасимпатический нейрон; 7 — гладкомышечные клетки тонкой кишки.
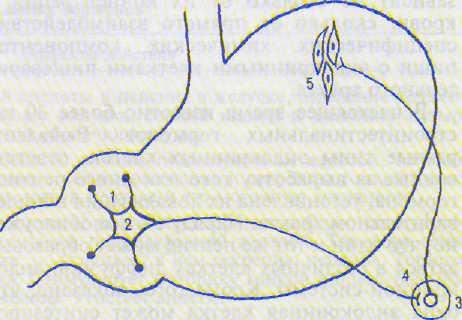
Рис. 14.2, Рефлекторная дуга периферического антрофундального тормозного рефлекса.
1 — механореиепторы антрального отдела желудка; 2 — сенсорный нейрон (клетка Догеля II типа); 3 — симпатический ганглий солнечного сплетения; 4 — ганглионарный симпатический нейрон; 5 — гладкомышечные клетки фундального отдела желудка.
тропанкреатохолецистодуоденальный комплекс), где особенно велика роль гастроин-тестинальных гормонов, высвобождаемых из эндокринных клеток желудочно-кишечного тракта и переносимых кровотоком к миоци-там, гландулоцитам и интрамуральным нейронам. Регуляторные пептиды могут также оказывать регулирующее влияние на секреторную и моторную функции паракринным путем (местно, не попадая в кровь). Поступая из оеток диффузной эндокринной системы в интерстиниальные пространства, они диффундируют через интерстициальную жидкость к близко расположенным клеткам-эффекторам и регулируют их деятельность.
Диффузная эндокринная система представляет собой совокупность рассеянных в слизистой оболочке желудочно-кишечного тракта и в поджелудочной железе специализированных эндокринных клеток, вырабатываю-
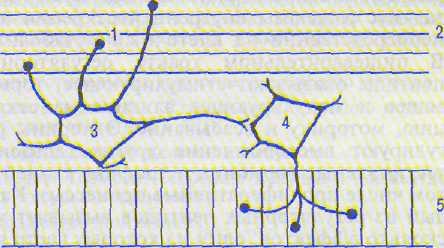
Рис. 14.3. Рефлекторная дуга местного моторного рефлекса желудка.
I — механореиепторы; 2 — продольный мышечный слой; 3, 4 — нейроны миэнтерального сплетения (3 — сенсорный, 4 — эфферентный холинергический); 5 — циркулярный мышечный слой.
щих биологически активные вещества. Продукты деятельности этой системы называют гастроинтестинальными гормонами, регуля-торными пептидами, энтеринами или пептид-гормонами. Высвобождение гастроин-тестинальных гормонов из эндокринных клеток происходит при участии блуждающего нерва, при действии на них других регулятор-ных пептидов, при механическом и химическом воздействиях пищи на клетки диффузной эндокринной системы. Регуляция образования пептид-гормонов в желудочно-кишечном тракте отличается от таковой эндокринных желез тем, что секреция гормонов
341
зависит не столько от их концентрации в крови, сколько от прямого взаимодействия специфических химических компонентов пищи с эндокринными клетками пищеварительного тракта.
В настоящее время известно более 30 га-строинтестинальных гормонов. Выявлены разные типы эндокринных клеток, ответственные за выработку того или иного пептид-гормона, установлена их локализация в пищеварительном тракте. Наряду с этим обнаружено, что один и тот же пептид может образовываться в различных клетках диффузной эндокринной системы. Кроме того, показано, что одна эндокринная клетка может синтезировать несколько регуляторных пептидов.
Гастроинтестинальные гормоны обладают широким спектром физиологической активности. Они не только участвуют в регуляции пищеварительных функций, но и вызывают общие эффекты, оказывая влияние на деятельность сердечно-сосудистой системы, ЦНС, регулируют обмен веществ. Многие пептиды пищеварительного тракта влияют на различные стороны метаболизма через гипоталамус и железы внутренней секреции или непосредственно на клеточный метаболизм. В пищеварительном тракте регуляторные пептиды оказывают стимулирующее, тормозящее и модулирующее влияние на секрецию, моторику и всасывание. Энтерины регулируют высвобождение других гастроин-тестинальных гормонов и трофику тканей (в том числе пролиферативные процессы). Каждый из регуляторных пептидов вызывает несколько эффектов, один из которых часто является основным (табл. 14.1).
Гастроинтестинальные гормоны относятся к группе «короткоживуших» химических веществ, период их полураспада составляет несколько минут. Вместе с тем вызываемые ими физиологические эффекты значительно продолжительнее. Особенностью регуляторных пептидов является способность одного и того же гормона по-разному влиять на моторику или секрецию разных отделов пищеварительного тракта, что обусловлено свойствами мембранных рецепторов различных органов-мишеней.
Концентрация многих гастроинтестиналь-ных гормонов в крови натощак изменяется в небольших пределах. Относительное постоянство содержания регуляторных пептидов в крови обеспечивается балансом поступления пептидов в кровоток и их ферментативного разрушения. Деградация пептид-гормонов происходит преимущественно в почках и печени и приводит к образованию более про-
стых пептидов — олигопептидов, обладающих большей или меньшей, иногда качественно иной активностью по сравнению с активностью исходных гастроинтестинальных гормонов. Дальнейший гидролиз пептидов приводит к полной потере их биологической активности.
Содержание некоторых регуляторных пептидов в крови в состоянии голода подвергается выраженным циклически повторяющимся колебаниям. Так, спонтанное повышение уровня мотилина в крови натошак обусловливает появление высоко амплитудных сокращений мышц желудка и кишечника, а его понижение приводит к прекращению моторной активности. Прием пиши вызывает повышение концентрации мотилина в крови и появление моторной активности мышц желудочно-кишечного тракта.
Прием пищи усиливает также процесс высвобождения и поступления в кровь гастри-на, секретина, холецистокинина, мотилина, ВИП, нейротензина и ряда других регуляторных пептидов. Характерными для энтеринов являются каскадный механизм выделения гормонов и запуск функций. Так, усиление секреторной активности обкладочных клеток под влиянием гастрина приводит к увеличению кислотности желудочного содержимого, поступление которого в двенадцатиперстную кишку вызывает высвобождение и поступление в кровь секретина и холецистокинина, которые не только стимулируют секреторную деятельность поджелудочной железы, но и оказывают тормозное влияние на секрецию кислого желудочного сока.
В. Фазы секреции пищеварительных желез. И.П.Павлов выделил две фазы секреции пищеварительных желез: сложнорефлекторную (ее называют также «мозговой») и нейрогу-моральную.
Первая (сложнорефлекторная) фаза секреции называется так потому, что она реализуется с помощью комплекса условных и безусловных рефлексов.
Во второй (нейрогуморальной) фазе секреции регуляция осуществляется посредством комплекса безусловных сокоотделительных рефлексов и гуморальных веществ.
В каудальном направлении ослабевает влияние ЦНС на пищеварительную систему и возрастает влияние гуморальных факторов и интраорганной нервной системы. Так, секреторная деятельность слюнных желез регулируется главным образом нервной системой.
В секреторной деятельности желез гастро-панкреатогепатодуоденального комплекса хорошо выражены оба механизма регуляции,
342
Таблица 14.1. Основные эффекты гастроинтестинальных гормонов
Гормон
Эффекты действия
Гастрин
Секретин
Холецистокинин (ХЦК)
Гастрин-рил изинг-пептид (бомбезин)
Вазоактивный интестинальный пептид (ВИП)
Гастрои нгибиру ющи й пептид (ГИП)
Мотилин
Панкреатический пептид (ПП)
Соматостатин Нейротензин
Энкефалин Энтероглюкагон
Глипентин (пептид YY) Вещество П
Серотонин Гнетам ин
Стимулирует секрецию соляной кислоты и пепсина в желудке, отделение поджелудочного сока. Усиливает моторику желудка, тонкой кишки и желчного пузыря. Вызывает гипертрофию слизистой оболочки желудка
Стимулирует секрецию бикарбонатов и воды поджелудочной железой, печенью, бруннеровыми железами. Усиливает желчеобразование и секрецию тонкой кишки. Тормозит секрецию соляной кислоты в желудке и его моторную активность
Усиливает моторику желчного пузыря, расслабляет сфинктер Одди, стимулирует желчевыделение, секрецию ферментов поджелудочной железой. Тормозит секрецию соляной кислоты в желудке, но усиливает в нем секрецию пепсина. Тормозит моторную деятельность желудка. Стимулирует моторику тонкой кишки. Вызывает гипертрофию поджелудочной железы. Угнетает аппетит
Стимулирует высвобождение гастрина и холецистокинина
Расслабляет гладкие мышцы кровеносных сосудов, желчного пузыря, желудка, тонкой кишки, сфинктеров. Тормозит действие ХЦК, секрецию желудка. Усиливает секрецию бикарбонатов поджелудочной железой, кишечную секрецию
Тормозит высвобождение гастрина, секрецию и моторику желудка. Усиливает глю-козозависимое высвобождение инсулина поджелудочной железой. Стимулирует кишечную секрецию
Усиливает моторику желудка и тонкой кишки. Стимулирует секрецию пепсина в желудке, секрецию тонкой кишки
Антагонист ХЦК. Тормозит секрецию ферментов и бикарбонатов поджелудочной железой. Расслабляет мышцы желчного пузыря. Усиливает моторику желудка и тонкой кишки. Стимулирует пролиферацию слизистой тонкой кишки, поджелудочной железы и печени Тормозит высвобождение гастроинтестинальных гормонов и секрецию желудка
Тормозит секрецию соляной кислоты в желудке. Стимулирует секрецию поджелудочной железы. Потенцирует действие секретина и ХЦК
Тормозит секрецию ферментов поджелудочной железой и желудком
Стимулирует мобилизацию углеводов. Тормозит секрецию желудка и поджелудочной железы. Ослабляет моторику желудка и кишечника
Тормозит секрецию желудка и поджелудочной железы
Усиливает слюноотделение, секрецию поджелудочной железы. Стимулирует моторику кишечника
Тормозит секрецию соляной кислоты в желудке. Стимулирует секрецию поджелудочной железы, желчевыделение, кишечную секрецию
Стимулирует секрецию желудка и поджелудочной железы, расширяет капилляры. Усиливает моторику желудка и кишечника
соответственно обе фазы секреции с преобладанием нейрогуморальной фазы.
В регуляции секреции кишечного сока ведущая роль принадлежит локальным механизмам, обусловливающим выделение секрета в месте действия химуса, обладающего свойствами механического и химического раздражителя.
Нервно-рефлекторные и гуморальные механизмы регуляции обеспечивают выделение всеми пищеварительными железами около 6—8 л секрета в сутки, большая часть которого подвергается обратному всасыванию в кишечнике.
14.2.4. ЗАКОНОМЕРНОСТИ ДЕЯТЕЛЬНОСТИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
А. Адаптация деятельности пищеварительных желез к различным пищевым веществам проявляется в соответствии объема, электролитного состава и спектра ферментов пищеварительных соков составу и количеству принятой пищи, что обеспечивает наиболее эффективный гидролиз пищевого субстрата. Основным фактором приспособления работы пищеварительных желез является химический состав пищи, действующий на рецепторные зоны пищеварительного тракта. Прием
343
любой пищи вызывает увеличение содержания всех ферментов в составе пищеварительного сока. Однако каждый вид пищи определяет доминирование в спектре гидролитической активности тех ферментов, которые обеспечивают переваривание соответствующего субстрата. В случае приема пищи с большим содержанием углеводов в пищеварительном соке максимально увеличивается концентрация амилазы, белков — лротеаз, а при еде жирной пищи — липаз. Различают два вида адаптации секреции пищеварительных желез: быструю (срочную) и медленную. 1. Быстрая адаптация состоит в приспособлении секреции ферментов и электролитов к определенному виду принятой пищи. Классическим примером приспособительных реакций являются кривые желудочной секреции на хлеб, мясо и молоко, полученные в лаборатории И.П.Павлова (см. рис. 14.8).
Наиболее эффективным фактором, стимулирующим отделение желудочного сока, является белковая пища животного происхождения (мясо), причем с максимальной концентрацией соляной кислоты. Наиболее слабым возбудителем кислой секреции желудочных желез является углеводистая пища (хлеб). Напротив, прием хлеба обусловливает наивысшую протеолитическую активность желудочного сока. Содержащиеся в хлебе углеводы и растительные белки оказывают выраженное стимулирующее влияние на деятельность главных (пепсиновых) клеток желудка. Такая реакция желудочных желез биологически целесообразна, поскольку только высокая концентрация протеаз в желудочном соке может обеспечить эффективный гидролиз трудно перевариваемых растительных белков. Этим же обстоятельством можно объяснить самую продолжительную секрецию желудочных желез на хлеб. Хлеб стимулирует отделение желудочного сока преимущественно в первой — сложнорефлекторной фазе секреции, а молоко — главным образом во второй, нейрогуморальной фазе секреции. При употреблении мяса хорошо выражены обе фазы желудочной секреции.
Секреторная деятельность поджелудочной железы быстро адаптируется не только к составу принятой пищи, но и к составу дуоденального содержимого. Чем выше кислотность желудочного содержимого, поступающего в двенадцатиперстную кишку, тем больше отделяется щелочного поджелудочного сока.
Наибольшее количество поджелудочного сока отделяется при употреблении хлеба, меньше — мяса и минимальное — молока. Наиболее щелочной сок выделяется на мясо, менее щелочной — на хлеб и молоко. Максимальная продолжительность секреции под-
желудочной железы отмечается при еде хлеба, а минимальная — при еде мяса (см. рис. 14.11).
2. Медленная адаптация деятельности пищеварительных желез заключается в постепенном и фиксируемом на значительное время приспособлении секреции к длительным рационам питания. Пищевой рацион определяет параметры секреции органов пищеварения, в том числе состав и свойства пищеварительных соков. Так, длительная мясная диета приводит к усилению отделения желудочного сока, повышению его кислотности и протеолитической активности. В зависимости от преобладания в пищевом рационе белков животного или растительного происхождения изменяется не только выработка протеолитических ферментов в желудке, но и их способность к эффективному гидролизу животных или растительных белков, названная соответственно зоо- и фитолити-ческой активностью (А.М.Уголев). Повышение зоолитической активности желудочного сока при длительном питании пищей, богатой животными белками, и фитолитической активности при употреблении растительной пиши обусловлено изменением кислотности и соотношения протеаз в секрете желудочных желез. При питании животными белками усиливается секреция поджелудочной железой трипсина, а при питании растительными белками — химотрипсина (А.М.Уголев).
Преимущественное углеводное питание увеличивает амилолитическую активность панкреатического сока, а большое количество жира в рационе стимулирует секрецию липазы. Способность поджелудочной железы адаптироваться к характеру пиши нарушается при недостатке в рационе белка.
Б. Конвейерный принцип организации деятельности пищеварительного тракта. И.П.Павлов сравнивал деятельность пищеварительного тракта с конвейерным химическим производством, обеспечивающим последовательную обработку пищи и продуктов ее расщепления с образованием по мере их продвижения все большего количества простых химических соединений. Б настоящее время работу пищеварительного конвейера рассматривают как совокупность последовательных этапов, обеспечивающих преемственность процессов физической и химической обработки пищи в ротовой полости, желудке, двенадцатиперстной кишке и проксимальном отделе тонкой кишки. Частично переваренная пища в ротовой полости передается в последующие отделы пищеварительного тракта, где подвергается дальнейшей механической и химической
344
обработке, обусловливающей гидролиз полимеров до стадии различных олигомеров, диме-ров и, наконец, мономеров.
Многократно повторяющиеся в различных отделах пищеварительного тракта процессы гидролитического расщепления пищевого субстрата в результате действия на него соответствующей группы ферментов можно рассматривать в качестве дублирующего механизма, повышающего надежность пищеварения.
У взрослого человека гидролиз жиров и липидов начинается в двенадцатиперстной кишке под действием панкреатической липазы и продолжается благодаря липолитичес-кой активности кишечного сока. В нормальных условиях основной гидролиз пищевых веществ завершается в проксимальном отделе тонкой кишки, тогда как дистальный его отдел компенсаторно включается в пищеварительный процесс, как правило, только при функциональной недостаточности проксимальной части тонкой кишки.
Г. Периодичность в деятельности пищеварительной системы. В деятельности пищеварительного тракта условно выделяют 3 основные группы ритмической активности: ба-зальные (секундные) ритмы с частотой от 3 ло 12 циклов в минуту; ритмы с частотой 7 — 14 циклов в сутки. Среди них особое место занимает околочасовой ритм внепищевари-тельной активности.
1. Базалъные (секундные) ритмы являются собственными ритмами пищеварительной системы и выражаются в способности гладких мышц спонтанно генерировать медленные электрические волны и потенциалы действия (ПД) и автоматически сокращаться. Генерация медленных электрических волн обусловлена спонтанным повышением проницаемости мембран гладкомышечных клеток для ионов натрия (фаза деполяризации) и последующим повышением проницаемости для ионов калия (фаза реполяризапии). В случае достижения критического уровня медленной деполяризации мембраны и повышения мембранной проницаемости для ионов кальция возникает группа (пачка) ПД, накладывающихся на медленные электрические волны (рис. 34.4). Генерация медленных электрических волн без ПД не приводит к сокращению гладких мышц желудочно-кишечного тракта. Появление пачек ПД на медленных электрических волнах обусловливает возникновение перистальтических сокращений, продолжительность которых соответствует периодам медленных волн, генерируемых гладкими мышцами в каждом отделе пищеварительного тракта.
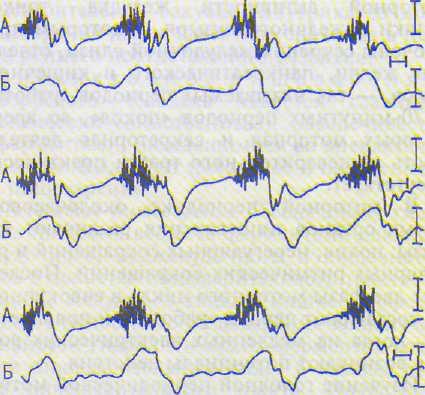
Рис. 14.4. Медленные электрические волны с пиковыми потенциалами (А) и сокращения (Б) гладких мышц желудка у кролика в условиях свободного поведения (объяснение в тексте).
Отметка времени 1 с; калибровочные сигналы: 0,5 мВ и 20 г.
Частота основного (базального) электрического ритма желудка у человека составляет 3 цикла/мин, ее задает водитель ритма, расположенный в области малой кривизны желудка вблизи кардии. Частота базального электрического ритма максимальна в двенадцатиперстной кишке (10—12 циклов/мин), что обусловлено наличием в ней, на уровне впадения желчного протока, водителя ритма, который задает частоту сокращений верхним отделам тонкой кишки. Частота генерации медленных волн и ритмических сокращений кишечника снижается в дистальном направлении и составляет у человека в нижних отделах тонкой кишки 8—6 циклов/мин и в толстой кишке — 6—1 циклов/мин. Уменьшение частоты базальных электрических ритмов по ходу кишечника в дистальном направлении получило название убывающего градиента автоматии. Базальные электрические ритмы пищеварительного тракта появляются на ранних этапах антенатального развития.
2. Околочасовой ритм пищеварительной системы — 90-минутный ритм внепищевари-тельной активности, открытый в лаборатории И.П.Павлова и получивший название голодной периодической деятельности пищеварительного тракта. Она наблюдается у человека и некоторых видов животных (собаки, кошки). Циклы голодной периодической деятельности состоят из 20-минутных периодов
345
моторной активности желудка, тонкой кишки и желчного пузыря, к которым приурочены секреция желудочной слизи, отделение желчи, панкреатического и кишечного соков, — так называемых периодов «работы» и 70-минутных периодов «покоя», во время которых моторная и секреторная деятельность пищеварительного тракта практически отсутствует.
В моторной периодике околочасового ритма органов пищеварения различают три фазы: покоя, нерегулярных сокращений и регулярных ритмических сокращений. Последние две фазы моторного цикла в совокупности составляют период «работы», и появляется он, когда на медленных электрических волнах возникают потенциалы действия.
Изучение голодной периодической моторной деятельности желудочно-кишечного тракта привело к представлению о мигрирующем моторном комплексе, под которым понимают постепенное перемещение моторной (миоэлектрической) активности от желудка и двенадцатиперстной кишки до терминальной части подвздошной кишки. Новый мигрирующий моторный комплекс возникает в желудке и двенадцатиперстной кишке, когда исчезает ритмическая активность подвздошной кишки.
Прием пищи устраняет мигрирующий моторный комплекс во всех отделах тонкой кишки. Вместо него возникает постоянная регулярная сократительная активность.
Голодная периодическая деятельность пищеварительного тракта является одним из проявлений цикличности в деятельности целого организма.
Ведущая роль в происхождении периодической деятельности пищеварительного тракта принадлежит ЦНС, которая определяет возникновение и последовательность перемещения миоэлектрической активности от двенадцатиперстной кишки до терминальной части подвздошной кишки.
В состоянии физиологического голода концентрация сахара в крови периодически изменяется. Минимальный уровень его содержания в крови, а также в слизистой оболочке желудочно-кишечного тракта отмечается в конце периодов «покоя». Эта информация воспринимается хеморецепторами сосудов и пищеварительного тракта и передается по сенсорным нейронам в гипоталамус. Снижение концентрации глюкозы в крови воспринимается и непосредственно гипоталамусом, где находятся центральные глюко-рецепторы. В результате этого гипоталами-ческий пищевой центр возбуждается и ока-
зывает наряду с активирующими восходящими влияниями на подкорковые структуры и кору большого мозга, ответственными за формирование пищевого поведения (поиск, пищедобывательное поведение и поедание пищи), нисходящие влияния, повышающие тонус ядер блуждающих нервов, что ведет к стимуляции деятельности желудочно-кишечного тракта.
При критически недостаточном уровне питательных веществ в крови гипоталамус оказывает активирующее влияние на ферментативные системы и механизмы обмена веществ, обеспечивающие возможность перехода организма на эндогенный тип питания за счет поступления в кровь питательных веществ из пищевых депо и других менее важных для сохранения жизни тканей.
Кроме того, при выделении в периоды «работы» в полость желудочно-кишечного тракта периодических секретов, содержащих большое количество белков и ферментов, происходят гидролиз этих белков и образование аминокислот, которые затем всасываются в кровь и усваиваются клетками и тканями организма.
Повышение уровня питательных веществ в крови воспринимается гипоталамусом, что приводит к активации симпатической нервной системы и неадренергических нейронов, в окончаниях которых выделяются АТФ и ВИП, обеспечивающие прекращение выхода питательных веществ из депо, а также моторной и секреторной деятельности желудочно-кишечного тракта. Наступает период «покоя». После того как ткани используют находящиеся в крови питательные вещества, циклические изменения моторной и секреторной активности пищеварительного тракта повторяются вновь.
Важную роль в формировании ритма играет интраорганная нервная система и гуморальные факторы, обусловливающие строго координированное распространение по тонкой кишке ритмических фаз мигрирующих моторных комплексов. Большое значение в самовозобновлении ритма периодической деятельности имеет поступление в двенадцатиперстную кишку периодических секретов, в том числе желчи, как внутриполостных стимуляторов сокращений кишечника.
Физиологическое значение голодной периодической деятельности пищеварительного тракта не ограничивается его участием в обмене веществ, обеспечивающим питание жизненно важных органов и тканей за счет собственных ресурсов организма. Моторная периодическая деятельность пищеваритель-
346
ного тракта обеспечивает эвакуаторную функцию по отношению к остаткам пищи; участие в поддержании гомеостазиса в результате выведения из крови шлаковых веществ (экскретов); препятствует распространению микрофлоры по тонкой кишке в проксимальном направлении, выполняя, таким образом, защитную функцию.
Периодическое выделение секретов пищеварительных желез в полость тонкой кишки в состоянии физиологического голода обеспечивает поддержание трофики ее слизистой оболочки, переваривание эндогенных белков и формирование ощущений чувства голода и насыщения.
14.2.5. ТИПЫ ПИЩЕВАРЕНИЯ
В зависимости от происхождения гидролаз различают три типа пищеварения: аутолити-ческое, симбионтное и собственное.
Аутолитическое пищеварение осуществляется гидролитическими ферментами, поступающими в пищеварительный тракт в составе пищевых продуктов. Роль его существенна на раннем этапе постнатального онтогенеза, когда недостаточно развито собственное пищеварение. В период молочного вскармливания аутолитический тип пищеварения обеспечивает у новорожденного створаживание материнского молока и гидролиз его компонентов за счет содержащихся в нем ферментов.
Симбионтное пищеварение реализуется благодаря действию гидролаз, синтезируемых симбионтами макроорганизма — бактериями и простейшими толстой кишки. У человека переваривание клетчатки в толстой кишке происходит под влиянием ферментов обли-гатной микрофлоры, в результате которого образуются вторичные нутриенты. Роль этого типа пишеварения у человека относительно невелика.
Собственное пищеварение. Человек и многие виды животных обладают собственным типом пищеварения, при котором макроорганизм использует для гидролиза пищевых веществ собственные ферменты, синтезированные пищеварительными железами.
В зависимости от локализации гидролитического процесса выделяют два типа пищеварения: внутриклеточное и внеклеточное.
Внутриклеточное пищеварение сводится к гидролизу мельчайших частиц пищевых веществ, поступивших в клетку путем эндоцитоза, при действии на них лизо-сомальных ферментов или в цитозоле, или в
пищеварительной вакуоли. Внутриклеточное пищеварение имеет известное значение в период раннего постнатального развития.
Внеклеточное пищеварение у взрослого человека является главным (практически единственным); обеспечивается ферментами, находящимися во внеклеточной среде. Выделяют полостное и пристеночное пищеварение.
Полостное пищеварение обеспечивает гидролиз пищевых веществ ферментами слюны, желудочного, поджелудочного и кишечного соков в полостях пищеварительного тракта — ротовой, желудке и тонкой кишке. Эффективность полостного пищеварения зависит от активности гидролитических ферментов секретов пищеварительных желез в каждом отделе пищеварительного тракта. Полостное пищеварение в тонкой кишке осуществляется как в жидкой фазе кишечного химуса, так и на границе фаз: на поверхности пищевых частиц, растительных волокон, сохраненных и разрушенных десквамированных энтероцитов. В результате полостного пищеварения образуются главным образом олигомеры. Последующий их гидролиз происходит в зоне, прилегающей к слизистой оболочке тонкой кишки.
Пристеночное пищеварение, открытое А.М.Уголевым, происходит на огромной поверхности тонкой кишки, образованной складками, ворсинками и микроворсинками слизистой оболочки, благодаря наличию гидролитических ферментов, адсорбированных из полости кишечника, а также встроенных в мембраны энтероцитов собственно кишечных ферментов.
Образующиеся в результате полостного гидролиза олигомеры вначале поступают в слой слизистых наложений, состоящий из мукоидного секрета бокаловидных клеток и фрагментов слущенного кишечного эпителия. Слой кишечной слизи удерживается гликокаликсом на пласте энтероцитов за счет высокой вязкости секрета. Он непрерывно обновляется. Слой слизистых наложений обладает более высокой гидролитической активностью, чем жидкое содержимое полости тонкой кишки. В нем сорбировано большое количество ферментов поджелудочного и кишечного соков, обеспечивающих частичный гидролиз олигомеров.
Богата ферментами также зона исчерченной каемки, образованная микроворсинками и мукополисахаридными нитями — гликокаликсом.
Между микроворсинками и на их апикальной поверхности располагается слой глико-
347
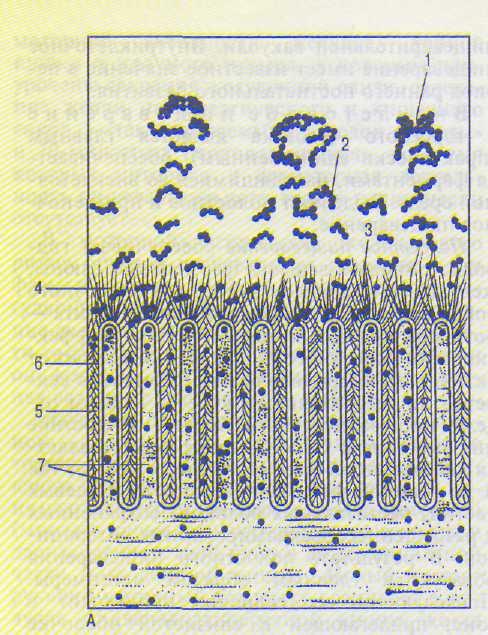
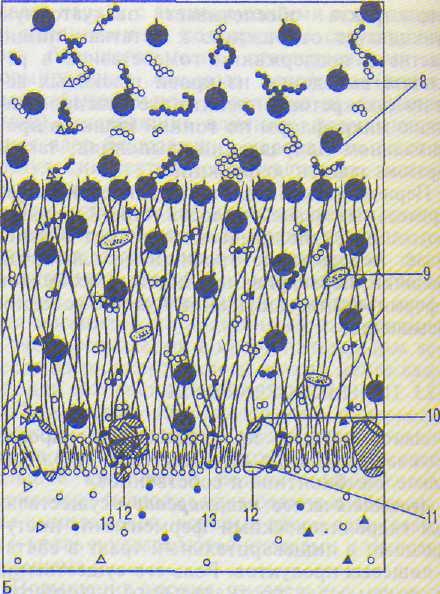
А — последовательная деполимеризация пищевых субстратов в полости и на поверхности тонкой кишки; Б — фрагмент липопротеиновой мембраны с адсорбированными и собственно кишечными ферментами.
1—3 — субстраты; 4 — апикальный гликокаликс; 5 — латеральный гликокаликс; 6 — мембрана; 7 — микроворсинки; 8 — панкреатические ферменты; 9 — неэнзиматические факторы; 10 — каталитические центры ферментов; 11 — мембранные ферменты; 12 — регуляторные центры ферментов; 13 — транспортная система мембраны.
каликса, в котором находятся панкреатические и кишечные ферменты, перешедшие из полости тонкой кишки, а также собственно кишечные ферменты, синтезируемые энтеро-цитами (рис. 14.5). Именно здесь продолжается гидролиз олигомеров, приводящий к образованию димеров, которые поступают на апикальные мембраны энтероцитов. Слой гликокаликса выполняет функцию своеобразного молекулярного сита, которое не только задерживает крупные молекулы, но и микроорганизмы. Поэтому пищеварение здесь протекает в стерильных условиях.
По А.М.Уголеву, окончательный гидролиз димеров, завершающийся образованием мономеров, осуществляют собственно кишечные ферменты, встроенные в апикальные мембраны энтероцитов, которые осуществляют мембранное пищеварение (см. рис. 14.5).
На апикальной поверхности каждого энте-роцита находится до 3 тыс. микроворсинок, а на 1 мм2 поверхности — до 50—200 млн, благодаря чему площадь мембран, на которых
совершается мембранное пищеварение, многократно возрастает. Гидролитические ферменты локализуются в мембранах этих микроворсинок. Эффективность мембранного пищеварения возрастает благодаря тому, что этот процесс сопряжен со следующим этапом — транспортом мономеров через энте-роцит в кровь или лимфу. Как правило, вблизи гидролитического фермента в апикальной мембране располагается транспортный механизм, который передает мономеры внутрь энтероцита (см. рис. 14.5).