

13.10. Регуляция системного артериального давления
13.10.1. ЦЕНТР КРОВООБРАЩЕНИЯ
Центром кровообращения считают совокупность нейронов, расположенных в различных отделах ЦНС и обеспечивающих приспособительные реакции сердечно-сосудистой системы в различных условиях жизнедеятельности организма (существует много других названий центра регуляции функции сосудов и сердца).
Локализация центра кровообращения была установлена с помощью метода перерезок и раздражения. Главная часть центра кровообращения, как и центра дыхания, находится в продолговатом мозге. Нейроны, регулирующие деятельность сердца и просвет сосудов, расположены также в среднем и спинном мозге, гипоталамусе, в коре большого мозга.
А. В спинном мозге совокупность симпатических нейронов, расположенных сегментар-но в боковых рогах, представляет собой конечное звено ЦНС, обеспечивающее передачу сигналов к эффекторам. Нейроны, регулирующие деятельность сердца, находятся в верхних грудных сегментах (Th,—Thnj), регулирующие тонус сосудов — в торако-люм-бальных сегментах (Cvm—LMI). Эти нейроны сохраняют самостоятельную активность и после перерезки спинного мозга в области нижних шейных или верхних грудных сегментов. Их импульсная активность приурочена к ритму сердца и колебаниям АД. О высоком уровне организации спинального отдела, связывающего центр кровообращения с периферическими эффекторами, свидетельствует факт быстрого восстановления АД после перерезки в эксперименте или в случае повреждения спинного мозга у человека. Сразу после повреждения спинного мозга АД падает до 60—70 мм рт.ст., затем оно в течение недели повышается до нормы, восстанавливаются при этом и сосудодвигательные рефлексы, возникающие на раздражение рецепторов кожи, мышц, соматических нервов, рецепторов растяжения сердечных камер, ме-ханорецепторов брюшины. АД восстанавливается у спинальных животных и после умеренной (до 25 %) кровопотери. Однако приспособительные реакции на изменения положения тела выражены крайне слабо. Следует учесть также, что в данных условиях в поддержании АД участвует и базальный тонус сосудов и деятельность сердца, которая регулируется блуждающими нервами. Центры последних локализованы в продолговатом мозге, и, естественно, этот регуляторный механизм не нарушен.
Б. В продолговатом мозге находятся центры блуждающих нервов. Нейроны, иннерви-рующие сердце, локализованы в обоюдном и дорсальном ядрах блуждающего (парасимпатического) нерва. С каждой стороны насчитывается около 500 нейронов. Сердечный парасимпатический (кардиоингибирующий) центр включает также ядра солитарного тракта (одиночного пути) и ретикулярные ядра — парамедианное, мелкоклеточное и
316
вентральное, к которым поступают афферентные импульсы от рецепторов сердца и сосудов. От нейронов этих ядер импульсы по коротким аксонам идут к нейронам ядер блуждающего нерва в продолговатом мозге. Аксоны нейронов ядер блуждающего нерва, не прерываясь, идут к сердцу и образуют синапсы с интраорганными холинергическими нейронами.
В 1871 г. Ф.В.Овсянников установил локализацию сосудодвигательного центра в продолговатом мозге в эксперименте с перерезкой ствола мозга: если перерезка производилась выше продолговатого мозга, АД не изменялось. В случае перерезки между продолговатым и спинным мозгом АД резко (до 60— 70 мм рт.ст.) уменьшалось. Позже было показано, что у бульбарного животного при изменении положения тела в пространстве возникают реакции сосудов, поддерживающие в них нормальное давление. У спинального животного эти реакции не развиваются. Название сосудодвигательный центр (продолговатый мозг) не соответствует реальной действительности, поскольку он является симпатическим центром, регулирующим не только тонус сосудов, но и деятельность сердца. Эта симпатическая часть центра кровообращения (сердечно-сосудистого центра), представляющая собой скопление нейронов ретикулярной формации, изучена недостаточно. Взаимоотношения нейронов симпатического центра значительно сложнее, чем парасимпатического.
-
Имеются прессорная и депрессорная его части, причем нейроны депрессорного отдела оказывают тормозное влияние на ней роны прессорной части центра кровообраще ния (см.рис.13.26), а их зоны расположения перекрывают друг друга.
-
Механизмы активации нейронов деп рессорного и прессорного отделов различны: депрессорные нейроны активируются аффе рентными импульсами от сосудистых бароре- цепторов (рецепторов растяжения, см. рис. 13.26, 1), а прессорные нейроны активируют ся афферентной импульсацией от сосудистых хеморецепторов и от экстерорецепторов (см. рис. 13.26,2). Аксоны прессорных нейронов продолговатого мозга посылают импульсы к симпатическим нейронам спинного мозга, иннервирующим и сердце (Th,—Thni), и со суды (Сущ—Lm). Медиатором прессорных и депрессорных нейронов продолговатого мозга является норадреналин. Медиатором преганглионарных симпатических нервных волокон, выходящих из спинного мозга, яв ляется ацетилхолин.
Прессорный отдел центра кровообращения находится в состоянии тонуса: в симпатических нервах постоянно идут нервные импульсы с частотой 1—3/с, при возбуждении — до 15/с. Именно поэтому при перерезке симпатических нервов сосуды расширяются. Однако деятельность сердца при блокаде симпатических нервов не изменяется. Это свидетельствует о том, что в покое такая им-пульсация не увеличивает частоту сердечных сокращений, но ведет к повышению тонуса сосудов. Тонус центра обеспечивается афферентными импульсами от баро- и хеморецепторов сосудов. Естественно, на все нейроны оказывают непосредственное влияние гуморальные факторы (гормоны, медиаторы, метаболиты). Кроме того, тонус центра кровообращения поддерживается и за счет спонтанной активности его нейронов, большинство из которых является нейронами ретикулярной формации ствола мозга. Они, как известно, обладают автоматией — способностью к спонтанной деполяризации, которая, достигнув критического уровня, обеспечивает возникновение потенциала действия. Нейроны центра кровообращения, как и нейроны других отделов ретикулярной формации, получают импульсы по коллатералям от всех специфических афферентных проводящих путей. Активность бульбарного отдела центра кровообращения регулируется гипоталамусом и корой большого мозга.
В. Гипоталамус, как и продолговатый мозг, содержит прессорные и депрессорные зоны, нейроны которых посылают аксоны к соответствующим центрам продолговатого мозга и регулируют их активность. На уровне гипоталамуса (промежуточный мозг) происходит интеграция соматических и вегетативных влияний нервной системы на организм — изменения соматической деятельности обеспечиваются соответствующими изменениями деятельности сердечно-сосудистой системы. Например, при физической нагрузке работа сердца увеличивается, происходит перераспределение крови в организме за счет сужения одних сосудов (кожи, пищеварительной системы) и расширения других сосудов (мышц, мозга, сердца), что ведет к увеличению кровотока в них, доставки кислорода, питательных веществ и удалению продуктов обмена. Раздражение ретикулярной формации в области среднего и промежуточного мозга (и особенно гипоталамуса) может оказывать на сердечно-сосудистую систему как стимулирующее, так и тормозное влияния. Изменяя область расположения электродов в гипоталамусе и параметры раздражения.
317
можно добиться локального сужения сосудов того или иного периферического русла, например почечного, мышечного или чревного.
При физическом и эмоциональном напряжениях возбуждение гипоталамуса сопровождается также выделением вазопрессина гипофизом, катехоламинов надпочечниками и ренина почками. Эти гормоны стимулируют деятельность сердечно-сосудистой системы. Афферентные импульсы гипоталамус получает от всех интеро- и экстерорецепторов.
Г. Влияние коры большого мозга на системное АД впервые выявили В.Я. Данилевский, Н.А- Миславский, В.М. Бехтерев. В опытах с раздражением теменной и лобной областей они обнаружили закономерные изменения кровяного давления вследствие изменения сосудистого тонуса. При раздражении различных отделов коры большого мозга чаше возникают прессорные эффекты, обычно в сочетании с увеличением ЧСС, реже наблюдаются падение давления и уменьшение ЧСС. Особенно сильное влияние на кровообращение оказывают моторная и премоторная зоны. Раздражение областей, вызывающих сокращение определенных мышц, вызывает локальное увеличение кровотока именно в этих мышцах, что является важной приспособительной реакцией, обеспечивающей соматическую деятельность вегетативными изменениями. Стимуляция передних отделов поясной извилины оказывает преимущественно депрессорный эффект, а воздействие на точки, расположенные около орбитальных областей островка височной коры, может сопровождаться как прессорными, так и деп-рессорными реакциями. Кора большого мозга реализует свое влияние на сердечнососудистую систему в обеспечении приспособительных реакций организма с помощью вегетативной нервной системы (условных, безусловных рефлексов) и гормональных механизмов. Таким образом, кора большого мозга и промежуточный мозг оказывают модулирующее влияние на бульбарныи отдел центра кровообращения, а при физической нагрузке и эмоциональном возбуждении влияние вышележащих отделов ЦНС сильно возрастает — наблюдается значительная стимуляция деятельности сердечно-сосудистой системы.
13.10.2. НЕСТАБИЛЬНОСТЬ АРТЕРИАЛЬНОГО ДАВЛЕНИЯ. ФАКТОРЫ, ВЛИЯЮЩИЕ НА ЕГО ВЕЛИЧИНУ
Величина АД является достаточно динамичной, она постоянно колеблется, особенно при физическом и эмоциональном напряже-
нии. Потребность в кислороде и питательных веществах отдельных органов в различных ситуациях сильно изменяется. Интенсивность кровотока в активно работающем органе нередко увеличивается в десятки раз, а работа сердца может возрастать только в 3— 5 раз. Поэтому кровеносные сосуды других органов должны сужаться, чтобы больше крови поступало в наиболее активно работающие органы. Это достигается с помощью взаимодействия местных и центральных ре-гуляторных механизмов. Однако даже в состоянии покоя организма поддерживать АД на постоянном уровне достаточно трудно, поскольку в аорте и крупных артериях содержится всего 6 % (0,35 л) от общего количества крови — это слишком мало по сравнению с общим количеством крови в сердечно-сосудистой системе (около 5 л). Кровь проходит по аорте очень быстро — со скоростью порядка 0,25 м/с, поэтому малейшая задержка поступления новых порций крови в аорту привела бы к катастрофическому падению АД. Но этого не происходит, что предотвращается совершенными механизмами регуляции АД. Постоянство (флюктуирующее) АД обеспечивает необходимый приток крови ко всем капиллярам организма, однако в капиллярах тоже очень мало крови — 6 % от общего количества. Кровообращение в организме существует ради пребывания крови в капиллярах, где проходят процессы газообмена и питательных веществ. Объемный кровоток прямо пропорционален количеству потребляемого кислорода. Если АД по какой-либо причине резко снизится, то функции всего организма сильно нарушатся. Поэтому в процессе эволюции выработались совершенные механизмы стабилизации АД. В некоторых органах (например, мозг, почки) более выражены специфические местные регуляторные механизмы, обеспечивающие постоянный кровоток.
Факторы, влияющие на АД: 1) работа сердца, 2) просвет сосудов, 3) объем циркулирующей крови (ОЦК) и 4) вязкость крови (при неизменной длине сосудов). Скорость изменения этих факторов различна. Работа сердца и просвет сосудов с помощью вегетативной нервной системы изменяются очень быстро — через несколько секунд. Гормональные влияния осуществляются медленнее. Исключение составляют адреналин и норадре-налин, вырабатываемые мозговым слоем надпочечников. Количество крови в организме и ее вязкость изменяются еще медленнее. Естественно, чем больше ОЦК, тем больше АД (ОЦК определяет величину среднего дав-
318
ления наполнения — давления в различных отделах сосудистого русла, которое устанавливается, когда сердце не работает). В зависимости от скорости включения и длительности действия все механизмы поддержания АД можно объединить в три группы: 1) механизмы быстрого реагирования; 2) механизмы небыстрого реагирования (средние по скорости включения и продолжительности действия); 3) механизмы медленного реагирования и длительного действия.
13.10.3. МЕХАНИЗМЫ БЫСТРОГО РЕАГИРОВАНИЯ
Механизмы быстрого реагирования — рефлекторная регуляция АД с помошью изменений работы сердца и тонуса (просвета) сосудов (рис. 13.25). Эти реакции срабатывают в течение нескольких секунд. В случае повышения АД работа сердца тормозится, тонус сосудов уменьшается — они расширяются. И то и другое ведет к снижению (нормализации) АД. Если же давление снижается, то деятельность сердца увеличивается, а сосуды сужаются, что ведет к увеличению — нормализации АД. При этом коррекция АД может быть избыточной. Например, при снижении АД работа сердца усилится, кровеносные сосуды сузятся так, что давление поднимется выше обычной нормы; в этом случае вновь включаются механизмы, понижающие АД, после нескольких флюктуации оно стабилизируется. Однако и в этом случае АД не остается абсолютно стабильным даже в покое, а флюктуирует, но со значительно меньшей амплитудой. Такой тип регуляции называется регуляцией по отклонению. Он реализуется согласно принципу обратной отрицательной связи. Включаются в реакцию и емкостные сосуды. В случае повышения АД тонус емкостных сосудов уменьшается, что ведет к задержке крови в венах, уменьшению притока крови к сердцу и уменьшению выброса крови сердцем. В случае снижения АД тонус емкостных сосудов возрастает, что ведет к увеличению возврата крови к сердцу и возрастанию выброса сердцем крови.
Рецепторы, воспринимающие изменения кровяного давления, — барорецепторы (точнее, рецепторы растяжения) рассеяны по всему кровеносному руслу, но имеются их скопления в дуге аорты и в области каротидного синуса (главные сосудистые рефлексогенные зоны), в сердце (предсердиях, желудочках, коронарных сосудах), легком, в стенках крупных грудных и шейных артерий.
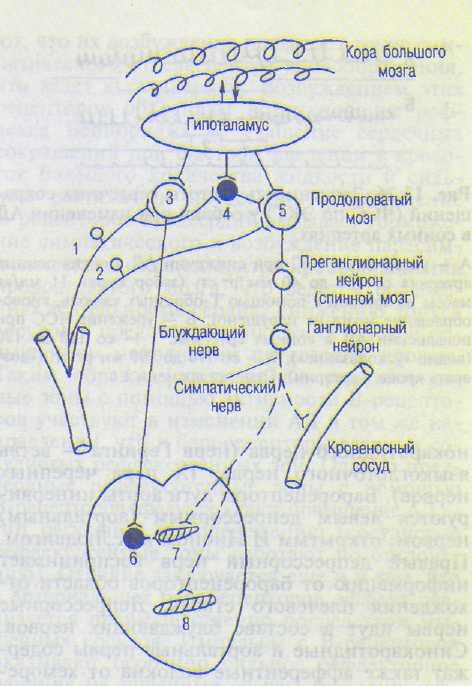
Рис. 13.25. Рефлекторная регуляция системного АД.
1 — рефлекс с барорецепторов; 2 — рефлекс с хеморецеп-торов; 3 — центр блуждающего (парасимпатического) нерва; 4 — депрессорная часть симпатического центра; 5 — прессорнал часть симпатического центра, — пост-ганглионарные нервные волокна; 6 — ганглионарный хо-линергический нейрон; 7 — синоатриальный узел; 8 — рабочий миокард.
В перечисленных участках имеются многочисленные барорецепторы, а в дуге аорты и каротидном синусе — баро- и хеморецепторы. Хотя принцип работы рефлексогенных зон одинаков, их значение в регуляции АД несколько различается.
А. Главные сосудистые рефлексогенные зоны расположены в начале напорного сосуда (дуга аорты) и в области каротидного синуса (участок, через который кровь течет в мозг) — эти зоны обеспечивают слежение за системным АД и снабжением кровью мозга. Отклонение параметров кровяного давления в области этих рефлексогенных зон означает изменение АД во всем организме, что воспринимается барорецепторами, и центр кровообращения вносит соответствующие коррекции. Чувствительные волокна от барорецепторов каротидного синуса идут в составе си-
319

Рис. 13.26. Зависимость частоты сердечных сокращений (ЧСС по ЭКГ) у собаки при изменении АД в сонных артериях.
А — увеличение ЧСС при снижении АД в обеих сонных артериях со 100 до 40 мм рт.ст. (забор крови 11 мл/кг массы из артерий с помощью Т-образных канюль, кровообращение мозга не нарушено); Б — урежение ЧСС при повышении АД в сонных артериях: 1 — со ПО до 170 (видно «ускользание»); 2 — со 3 70 до 190 мм рт.ст. («возврат» крови в артерии). Отметка времени ! с.
нокаротидного нерва (нерв Геринга — ветвь языкоглоточного нерва, IX пара черепных нервов). Барорецепторы дуги аорты иннерви-руются левым депрессорньш (аортальным) нервом, открытым И.Ционом и К.Людвигом. Правый депрессорный нерв воспринимает информацию от барорецепторов области от-хождения плечевого ствола. Депрессорные нервы идут в составе блуждающих нервов. Синокаротидные и аортальные нервы содержат также афферентные волокна от хеморе-цепторов, расположенных соответственно в каротидных (около области разветвления общей сонной артерии) и аортальных тельцах (дуга аорты).
При снижении АД барорецепторы рефлексогенных зон возбуждаются меньше. Это означает, что меньше поступает импульсов от дуги аорты и синокаротидной области в центр кровообращения. В результате нейроны блуждающего нерва меньше возбуждаются и к сердцу по эфферентным волокнам поступает меньше импульсов, тормозящих работу сердца, поэтому частота и сила его сокращений возрастают (рис. 13.26, А). Одновременно меньше импульсов поступает к депрессорным нейронам симпатического отдела центра кровообращения в продолговатом мозге и вследствие этого его возбуждение ослабевает, меньше угнетаются прессор-ные нейроны, а значит, они посылают больше импульсов к сердечным (Th|—Thjn) и сосудистым (Cvm—Ljjj) симпатическим центрам спинного мозга. Это ведет к дополнительному усилению сердечной деятельности и сужению кровеносных сосудов. Сужаются при этом венулы и мелкие вены, что увеличивает возврат крови к сердцу и ведет к усилению его деятельности. В результате согласованной деятельности симпатического и парасимпа-
тического отделов центра кровообращения АД повышается (нормализуется).
При повышении АД увеличивается импульсация от барорецепторов в центр кровообращения, что оказывает депрессорное действие — снижение АД. Наибольший прирост импульсации наблюдается при увеличении АД от 80 до 180 мм рт.ст. Импульсация носит залпообразный характер, приуроченный к пульсации сосудов, — на вершине пульсовой волны импульсация наибольшая. Чем выше давление, тем больше прирост импульсации (нарастание в геометрической прогрессии). Например, в одиночном афферентном волокне в диапазоне прироста давления 130—140 мм рт.ст. прирост импульсации составляет 5 имп/с, а в диапазоне 180— 190 мм рт.ст. прирост достигает 25 имп/с, частота в течение каждой секунды становится больше на 25 импульсов. При этом одна группа рецепторов воспринимает давление от 80 до 120 мм рт.ст., другая группа воспринимает давление ниже 80, третья — выше 120 мм рт.ст. Естественно, в каждой из этих групп есть и подгруппы. Поскольку артериальное давление у человека очень часто изменяется, то различные группы рецепторов сменяют друг друга в работе, что обеспечивает поступление информации в ЦНС о величине АД в широком диапазоне. Есть рецепторы, которые воспринимают в основном пульсовое давление, другие — в основном постоянную составляющую давления. Снижение повышенного АД до уровня нормы осуществляется с помощью увеличения поступления числа импульсов от рефлексогенных зон в центр кровообращения. Усиление возбуждения нейронов блуждающего нерва (увеличение его тонуса) ведет к угнетению сердечной деятельности (рис. 13.26, Б), а усиление возбуждения депрессорной части симпатического центра ведет к большему угнетению прес-сориого отдела симпатического центра и к расширению резистивных и емкостных сосудов организма. В результате угнетения работы сердца и расширения сосудов давление понижается. Оно дополнительно уменьшается еще и потому, что задержка крови в расширенных емкостных сосудах ведет к уменьшению поступления крови к сердцу и, естественно, к уменьшению систолического выброса крови.
Однако при значительном (180 мм рт.ст. и выше) увеличении АД через несколько дней наступает адаптация сосудистых барорецепторов, работающих в этом диапазоне, частота импульсации снижается и приближается к нормальной. Поскольку афферентная им-
320
; пульсация нормализуется, в ЦНС это высо-\ кое АД оценивается как нормальное, рефлек-', торные механизмы поддерживают его тако-; вым — порочный круг.
Следует отметить, что увеличение аффе-' рентной импульсации от барорецепторов ве-' дет к торможению некоторых отделов ЦНС, t в результате чего, например, дыхание стано-' вится более поверхностным, снижается мы-. шечный тонус, наблюдается тенденция к синхронизации ЗЭГ, как это имеет место , при засыпании. В эксперименте у бодрствующих животных при сильном растяжении области каротидного синуса наблюдается снижение двигательной активности; иногда они даже засыпают.
Возбуждение хеморецепторов аортальной и синокаротидной рефлексогенных зон возникает при уменьшении напряжения О2, увеличении напряжения СО2 и концентрации водородных ионов, т.е, при гипоксии, гиперкапнии и ацидозе. Импульсы от хеморецепторов поступают по тем же нервам, что и от барорецепторов, в продолговатый мозг, но непосредственно к нейронам прессорного отдела симпатического центра, возбуждение которого вызывает сужение сосудов, усиление и ускорение сердечных сокращений и как следствие — повышение АД. В результате кровь быстрее поступает к легким, углекислый газ обменивается на кислород. Хеморецепторы имеются и в других сосудистых областях (селезенка, почки, мозг). Изменения деятельности сердечно-сосудистой системы способствуют устранению отклонений от нормы газового состава крови. Однако эффект невелик, так как увеличение АД осуществляется главным образом за счет сужения сосудов и лишь частично — в результате стимуляции деятельности сердца. Такой же эффект наблюдается и при нарушении кровоснабжения мозга любого происхождения (недостаточности деятельности сердца, нарушении мозгового кровообращения). Если степень нарушения кровоснабжения мозга выражена сильно, АД может повыситься до 200 мм рт.ст. и больше.
Б. Сердечные рефлексогенные зоны. В предсердиях, особенно в области впадения полых и легочных вен, имеются рецепторы растяжения (Аи В), импульсы от которых поступают в центр кровообращения по блуждающим нервам. А-рецепторы возбуждаются при сокращении предсердий, В-рецепторы — при пассивном растяжении предсердий во время наполнения их кровью, в основном в конце диастолы. Физиологическое значение А-рецепторов изучено недостаточно. Полага-
ют, что их возбуждение повышает тонус симпатического отдела центра кровообращения, что ведет к тахикардии. Возбуждением этих рецепторов объясняют возникновение рефлекса Бейнбриджа — учащение сердечных сокращений при быстром введении в кровоток большого количества жидкости и сильном растяжении предсердий. Возбуждение В-рецепторов предсердий вызывает торможение симпатического и возбуждение парасимпатического отделов центра кровообращения продолговатого мозга, что ведет к ослаблению деятельности сердца, снижению тонуса сосудов и предотвращению роста АД при повышенном наполнении предсердий кровью. Таким образом, предсердные рефлексогенные зоны с помощью активности В-рецепторов участвуют в изменении АД в том же направлении, что и барорецепторы главных сосудистых рефлексогенных зон. Однако предсердные рефлексогенные зоны с помощью В-рецепторов предупреждают повышение АД — тип регуляции по опережению (сосудистые рефлексогенные зоны работают по отклонению). Отличие заключается еще и в том, что с артериальных барорецепторов эффект реализуется преимущественно посредством мышечных сосудов. Импульсы с В-рецепторов оказывают особенно сильное рефлекторное влияние на почечные сосуды (усиление афферентной импульсации ведет к сужению сосудов почек, ослабление — к расширению их). Однако изменение кровенаполнения почек непосредственно на системном давлении сильно не сказывается, но может привести к изменению интенсивности образования мочи и как следствие — к изменению объема циркулирующей крови и АД. В стенках желудочков также имеется некоторое количество рецепторов растяжения, импульса-ция от них поступает в центр кровообращения по блуждающим нервам и возбуждает их центры, что поддерживает тоническое отрицательное хронотропное влияние на деятельность сердца.
В. Легочная рефлексогенная зона. Барорецепторы (механорецепторы) этой зоны локализуются в артериях малого круга кровообращения. Повышение давления в сосудах легких закономерно ведет к урежению сокращений сердца, к падению АД в большом круге кровообращения и увеличению кровенаполнения селезенки (рефлекс Парина). Попадание в сосуды легких (в патологических случаях) пузырьков воздуха, жировых эмболов, вызывающих раздражение механорецепторов сосудов малого круга кровообращения, настолько сильно угнетает сердечную деятель-
321
ность, что может привести к летальному исходу — нормальная физиологическая реакцию переходит в случае чрезмерного ее проявления в патологическую.
13.10.4. МЕХАНИЗМЫ НЕБЫСТРОГО И МЕДЛЕННОГО РЕАГИРОВАНИЯ
А. Механизмы небыстрого реагирования —
это средние по скорости развития реакции (минуты — десятки минут), участвующие в регуляции АД. Они включают четыре основных механизма.
1. Изменение скорости транскапиллярного перехода жидкости, что может осуществляться в течение 5—10 мин в значительных количествах. Повышение АД ведет к увеличению фильтрационного давления в капиллярах большого круга кровообращения и, естественно, к увеличению выхода жидкости в межклеточные пространства и нормализации АД. Увеличению выхода жидкости способствует также повышение кровотока в капиллярах, которое является следствием рефлекторного расширения сосудов при росте АД. При снижении АД фильтрационное давление в капиллярах уменьшается, вследствие чего повышается реабсорбция жидкости из тканей в капилляры, в результате АД возрастает. Данный механизм регуляции АД работает постоянно, особенно сильно он проявляется после кровопотери. Если величина потери крови не превышает 15 мл/кг, то среднее АД практически не изменяется, при большей кровопо-тере оно резко падает. Реабсорбция жидкости из тканей в совокупности с другими регуля-торными механизмами ведет к повышению (нормализации) АД. Уже через 15—30 мин после потери 500 мл крови 80—100 % плазмы восполняется за счет межклеточной жидкости. При большей потере объем плазмы нормализуется через 12—72 ч, при этом, естест-
венно, включаются механизмы и длительного действия.
-
Увеличение или уменьшение объема депо нированной крови, количество которой со ставляет 40—50 % от общего объема крови. Функцию депо выполняют селезенка (около 0,5 л крови), сосудистые сплетения кожи (около 1 л крови), где кровь течет в 10—20 раз медленнее, печень и легкие, причем в селе зенке кровь сгущается и содержит до 20 % эритроцитов всей крови организма. Кровь из депо может мобилизоваться и включаться в общий кровоток в течение нескольких минут. Это происходит при возбуждении симпати- ко-адреналовой системы, например, при фи зическом и эмоциональном напряжении, при кровопотере.
-
Изменение миогенного тонуса — см. раз дел 13.9.2.
-
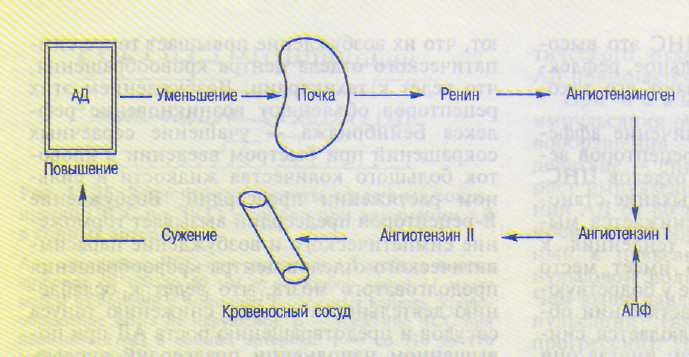
Вследствие изменения количества выра ботки ангиотензина (рис. 13.27).
В нормальных физиологических условиях после падения системного АД максимальный эффект увеличения выработки ренина, активирующего ангиотензиноген, развивается в течение 20 мин. Ангиотензин, возбуждая симпатико-адреналовую систему, усиливает также работу сердца, увеличивает венозный приток крови к сердцу вследствие сужения вен — все это ведет к повышению АД. Следует отметить, что ухудшение кровоснабжения почки, например сужение приносящей артерии в результате патологического процесса, также стимулирует выброс ренина, вследствие чего может развиться гипертензия. Ангиотензин активирует также выработку альдос-терона, что может привести к повышению АД, но это уже механизм более длительного действия.
Б. Механизмы медленного реагирования — это регуляция системного АД с помощью изменения количества выводимой из организма воды. При увеличении количества воды в ор-
322
ганизме, несмотря на переход части ее из кровеносного русла в ткани, АД возрастает по двум причинам: а) из-за непосредственного влияния количества жидкости в сосудах: чем больше крови, тем больше давление в сосудах — возрастает давление наполнения; б) при накоплении жидкости в кровеносном русле возрастает наполнение емкостных сосудов (венул и мелких вен), что ведет к увеличению венозного возврата крови к сердцу и. естественно, к увеличению выброса крови в артериальную систему — АД повышается. При уменьшении количества жидкости в организме АД уменьшается. Количество выводимой из организма воды определяется фильтрационным давлением в почечных клубочках и меняется с помощью гормонов.
-
Фильтрационное давление в почечных клубочках определяет количество первичной мочи. При увеличении АД растет, естествен но, и фильтрационное давление, что ведет к возрастанию объема фильтрата (первичной мочи) в почечных клубочках. Кроме того, скорость движения жидкости в канальцах почки становится больше, в результате чего меньше реабсорбируется воды из канальцев — диурез возрастает, ОЦК уменьшается, что ведет к снижению АД. В случае уменьшения АД развиваются противоположные эффекты. Следует также заметить, что обычно повы шенное потребление воды не сопровождается существенным подъемом кровяного давле ния, так как избыточная вода выводится из организма почками. Даже если систематичес ки потреблять повышенное количество воды, она не задерживается в организме — ее боль ше выводится. При этом устанавливается равновесие между приходом и расходом жид кости на новом, более высоком уровне. Од нако регуляция выведения воды из организма за счет изменения фильтрационного давле ния играет второстепенную роль, так как ми- огенный механизм регуляции почечного кро вотока стабилизирует его в пределах измене ния системного АД от 80 до 180 мм рт.ст. Главную роль играют гормоны.
-
Гормональная регуляция. Антидиурети ческий гормон (АДГ) участвует в регуляции АД посредством изменения количества выво димой из организма воды лишь в случае зна чительного его падения. При увеличении объема крови импульсация от рецепторов предсердий и крупных вен (вследствие их растяжения — волюморецепторы) возрастает, в результате через 10—20 мин выделение АДГ снижается. Это приводит к увеличению вы деления жидкости почками и к постепенному снижению АД. Волюморецепторы располо-
жены в предсердиях, преимущественно в области впадения полых и легочных вен. При падении кровяного давления происходят обратные процессы: выброс АДГ возрастает и выделение жидкости уменьшается, что ведет к медленному повышению АД.
Альдостерон участвует в регуляции системного АД, во-первых, за счет повышения тонуса симпатической нервной системы и повышения возбудимости гладких мыши сосудов к вазоконстрикторным веществам, в частности к ангиотензину, адреналину, вызывающим сужение сосудов (по-видимому, повышается активность а-адренорецепто-ров). В свою очередь ангиотензин оказывает сильное стимулирующее влияние на выработку альдостерона — так функционирует ренин-ангиотензин-альдостероновая система. Во-вторых, альдостерон участвует в регуляции АД за счет изменения объема диуреза. При уменьшении ОЦК и снижении АД альдостерона вырабатывается больше, он увеличивает реабсорбцию Na+ в канальцах почки, повышая тем самым его содержание в организме. Вместе с Na1" задерживается и вода согласно закону осмоса, что ведет к повышению (нормализации) АД. При увеличении АД выработка альдостерона уменьшается, Na+ выводится из организма в больших количествах, что ведет к увеличению выведения воды из организма и снижению АД. Эффекты альдостерона начинают проявляться через несколько часов после отклонения АД от нормы и достигают максимума через несколько дней.
Натрийуретические гормоны являются антагонистами альдостерона в регуляции содержания Na+ в организме: они способствуют выведению Na+. Этим гормонам, секретиру-ющимся в миокарде, почках, мозге, посвящено огромное количество работ, они представляют собой пептиды. Атриопептид вырабатывается кардиомиоцитами в основном в предсердиях, частично в желудочках. При увеличении растяжения предсердий продукция гормона возрастает. Это наблюдается при увеличении объема циркулирующей жидкости в организме и кровяного давления. Повышение выведения Na+ с мочой ведет к увеличению выведения воды, уменьшению (нормализации) АД. Снижению АД способствует также сосудорасширяющее действие этих гормонов, что осуществляется с помощью ингибирования Са-каналов сосудистых мио-цитов. Атриопептид увеличивает мочеобразо-вание также посредством расширения сосудов почки и увеличения фильтрации в почечных клубочках. При уменьшении объема
323
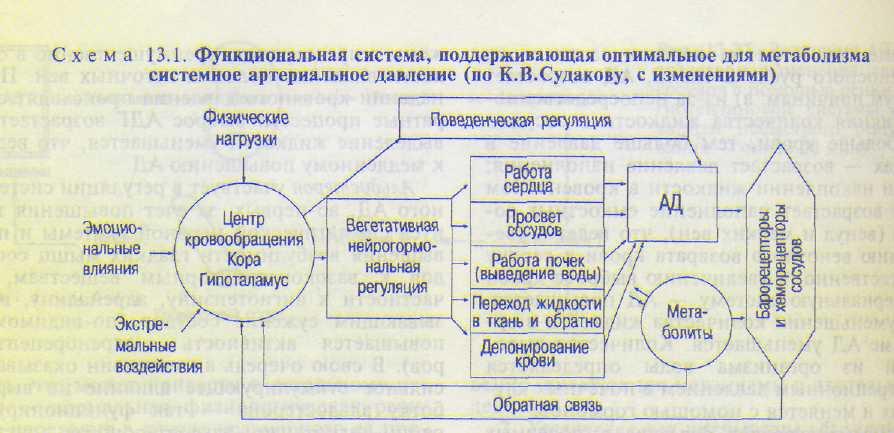
Важно отметить, что все рассмотренные механизмы регуляции АД взаимодействуют между собой, дополняя друг друга в случае как повышения, так и понижения АД. Например, при потерях крови до 25 % взаимодействие всех регуляторных механизмов приводит к повышению (нормализации) АД и стабилизации его. Однако потеря большего количества крови опасна для жизни, а при быстрой потере 40—50 % крови АД резко падает вплоть до нуля и наступает смерть. Схема функциональной системы, поддерживающей оптимальный для метаболизма уровень АД, представлена на схеме 13.1.