

14
Значительная часть гидролаз представлена пищеварительными ферментами, с помощью которых полимеры, поступающие с пищей в организмы животных и человека, расщепляются до мономеров и реутилизируются для построения новых биополимеров по собственным наследственным программам.
Гидролиз полимеров помимо его значения для реутилизации важнейших мономеров, играет в живой природе еще две важные роли.
Во-первых, это уничтожение внутри клетки биополимеров, которые, выполнив свою функцию, становятся ненужными на последующем этапе жизнедеятельности. Известно, например, что у прокариот время жизни отдельных молекул мРНК невелико – выполнив свою функцию в производстве необходимых клетке белков, они гидролизуются до мономеров и повторно используются для создания новых молекул нуклеиновых кислот, необходимых для следующего этапа жизни.
Во-вторых, это осуществление так называемого процессинга белков и РНК. Многие белки и РНК синтезируются не в зрелом виде, а в виде предшественников, содержащих дополнительные фрагменты полипептидных и полинуклеотидных цепей. Эти фрагменты удаляются с помощью специальных гидролаз, и такая обработка предшественников с образованием зрелых молекул называется процессингом.
Гидролазы находят применение и в пищевых технологиях. В хлебопекарном и пивоваренном производствах используются амилазы для улучшения качества теста и осоложения крахмала. Протеиназы и целлюлазы оказываются полезными реагентами в виноделии и приготовлении соков. Протеиназы ускоряют процесс мягчения мяса, улучшающего консистенцию, а также применяются при производстве сыров.
4.1.Глюкозидазы
4.1.1.Выделение α– и β– амилаз солода.
Амилазы гидролизуют различные соединения: амилозу, амилопектин, гликоген, олигосахариды и родственные им вещества, построенные из остатков α–D–глю- копиранозы и содержащие в молекулах 1,4 –1,6 гликозидные связи.
Активные центры амилаз образованы радикалами гистидина, аспарагиновой и глутаминовой кислот, а также тирозина. Последнему приписывают функцию связывания субстрата, а первым трем – каталитическую функцию. Предполагают следующий механизм распада гликозидных связей 1,4 – 1,6 при участии амилаз (см. рисунок 3 (I стадия)).
На второй стадии в активный центр фермента входит молекула воды, присоединения ОН групп, которой обеспечивает распад возникшей сложно–эфирной связи и высвобождение радикала дикарбоновой аминокислоты; протон воды присоединяется к депротонированному на 1–ой стадии радикалу гистидина (см. рисунок 4 (II стадия)).
15
В результате восстанавливается структура активного центра фермента, и он готов к осуществлению следующего каталитического акта в гидролизе гликозидной связи.
16

17
Радикал дикарбоновой аминокислоты
активного центра
|
|
|
|
O |
|
|
|
|
OH |
|
С |
OH |
|
|
H2C |
|
O- H2C |
|
||
|
|
|
|
|
||
|
H |
|
O H |
H |
O H |
|
|
H |
|
|
H |
|
|
В цепь |
OH |
H |
|
OH |
H |
В цепь |
|
О |
|
|
О |
О |
|
|
H |
OH |
|
H |
OH |
|
|
|
HN |
|
NH+ |
|
|
СН2
Протонированный радикал гистидина
активного центра
|
H2C |
OH |
|
С |
O |
H2C |
OH |
|
|
|
|
||||
|
H |
|
O |
O |
H |
|
O H |
|
H |
|
|
|
|
H |
|
В цепь |
OH |
H |
|
|
|
OH |
H |
О |
|
|
H |
H |
|
О В цепь |
|
|
|
|
|
||||
|
H |
OH |
|
|
|
H |
OH |
|
|
|
|
|
|
Олигосахарид |
|
|
|
HN |
|
N |
|
|
|
СН2
Рисунок 3 – Механизм действия амилаз (1-я стадия)

18
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
С |
|
H2C |
OH |
С |
O |
H2C |
OH |
O- |
|
|
|
|
||||
|
H |
O |
O |
|
H |
O H |
|
|
H |
|
|
|
H |
|
|
В цепь |
OH |
H |
|
В цепь О |
OH |
H |
|
О |
|
H |
|
H |
|
||
|
H |
OH |
|
+ НО --- Н |
H |
OH |
|
|
|
|
|
|
Олигосахарид |
NH+ |
|
|
HN |
|
N |
|
|
HN |
|
СН2 СН2
Рисунок 4 – Механизм действия амилаз (2-я стадия)
Предполагают, что не только амилазы, но и все другие гликозидазы действуют в соответствии с приведенным ниже механизмом.
Известны три вида амилаз, различающихся по распространению в природе, по ряду свойств и главным образом по конечным продуктам ферментативного действия.
α–Амилаза (α–1,4–глюкан–4–глюканогидролаза) – фермент гидролизует внутренние α–1,4– глюкозидные связи в субстратах, приводя в конечном итоге к образованию мальтозы, некоторого количества глюкозы и различных низкомолекулярных олигосахаридов с разветвленной цепью.
β–Амилаза (α–1,4–глюканмальтогидролаза) и глюкоамилаза (α–1,4–глюкан глюкогидролаза) в отличие от α–амилазы гидролизуют наружные нередуцирующие концы цепей полисахаридов, последовательно отщепляя соответственно мальтозные и глюкозные остатки. Действие β–амилазы и глюкоамилазы прекращается, как правило, вблизи первой точки ветвления полисахарида, образованного α–1,6– глюкозидной связью. Поэтому, помимо мальтозы и глюкозы, под действием β–амилаз и глю-
коамилаз образуются декстрины (олигосахариды) высокой молекулярной массы. Ход работы. 20 г измельченного солода помещают в мерную колбу на 100 мл,
доводят до метки дистиллированной водой и переносят в фарфоровый стакан. Затем массу размешивают 15 минут, оставляют на 2 часа в холодильнике, перемешивают,
19
отжимают через ткань и центрифугируют. Вытяжка из солода содержит α–и β– амилазы.
Выделение α–амилазы
10 мл солодовой вытяжки нагревают в пробирке при 70 °С в течение 15 минут, после чего раствор охлаждают и используют для исследования активности фермента. β–амилаза при этой температуре инактивируется.
Выделение β–амилазы
15 мл солодовой вытяжки вносят в стакан, добавляют 3 мл 0,1 н. раствора соляной кислоты и 12 мл воды до общего объема 30 мл (рН при этом должно быть 3,3) и оставляют в холодильнике на15 мин. Затем к раствору добавляют 6 мл 0,15 М раствора фосфорнокислого натрия (двухзамещенного) с целью доведения рН среды до 6,0. Раствор берут для определения осахаривающей способности β–амилазы, так
как α– амилаза в кислой среде инактивирована.
4.1.2. Определение декстринирующей способности α–амилазы
Принцип действия. При действии на крахмал α–амилазы: образуется большое количество декстринов разного молекулярного веса, по–разному окрашивающихся йодом – амилодекстрины, эритродекстрины, ахродекстрины, мальтодекстрины.
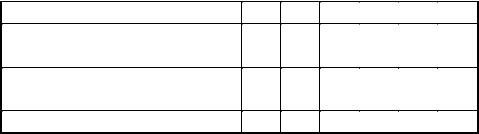
Ход работы. В шесть пробирок вносят по 5 мл 2 %–ного раствора растворимого крахмала. В каждую пробирку добавляют по 1 мл буферного раствора с рН 5,6. Раствор фермента и воду вносят в количествах, указанных в таблице 2:
Растворы в пробирках хорошо перемешивают и ставят в термостат при 40 °С. Глубину гидролиза крахмала контролируют по четвертой пробирке. Для этого из нее стеклянной палочкой периодически отбирают пробы на белую кафельную плитку, добавляя к ней раствор Люголя. Первый отбор пробы делают через пять минут после термостатирования. Пробирки выдерживают в термостате до тех пор, пока проба из четвертой пробирки не окрасится в красно–бурый цвет, что свидетельствует об образовании в этой пробирке эритродекстринов.
Таблица 2 – Основные параметры опыта и его результаты
|
|
|
|
|
|
|
|
№ пробирки |
1 |
2 |
3 |
4 |
5 |
6 |
|
|
|
|
|
|
|
|
|
Количество |
– |
0,2 |
0,4 |
0,6 |
0,8 |
1,0 |
|
фермента, мл |
|||||||
|
|
|
|
|
|
||
Количество |
4 |
3,8 |
3,6 |
3,4 |
3,2 |
3,0 |
|
воды, мл |
|||||||
|
|
|
|
|
|
||
Образовавшиеся декстрины |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20
После этого в каждую пробирку добавляют по 2–3 капли раствора йода и 20 %–ного раствора соляной кислоты для остановки гидролиза крахмала. Содержимое пробирок окрашивается в различные цвета: от синего в первой пробирке с крахмалом до желтого – в последней, через фиолетовый и красно–бурый.
Анализируя окраску с йодом, в таблицу 2 следует записать, до каких декстринов прошел гидролиз крахмала в каждой пробирке в зависимости от количества фермента.
Материалы и реактивы. Солод; 0,1 н раствор HCI; 0,15 М раствор фосфата натрия (двузамещенный); 2 % – ный раствор крахмала; 0,1 М раствор фосфатного буфера, рН 5,6 (см. Приложение 1, п.3); 20 %–ный раствор НСI; раствор Люголя.
Оборудование. Термостат; холодильник; центрифуга; кафельные плитки; стеклянные палочки; пипетки на 5,10,15 мл; фарфоровые стаканы; водяная баня.
4.1.3. Влияние температуры на осахаривающую способность β–амилазы
Каталитическая активность ферментов значительно зависит от температуры. При низкой температуре ( 0 °С, 4 °С) ферментативные реакции протекают с малой скоростью, а при замораживании могут и не происходить. Температурный оптимум многих ферментов лежит в пределах от 40 °С до 50 °С. При нагревании свыше 60–70 °С ферменты инактивируются необратимо, меняя свое строение, что приводит к потере каталитической активности.
Работа выполняется коллективно всей группой, каждый студент получает индивидуальное задание при какой температуре проводить исследование. Опыты ставят
при температуре 4 °С (холодильник), 20 °С (комнатная), 30 °С, 40 °С, 50 °С, 60 °С, 70 °С (водяные бани или термостат).
Чтобы получить правильные результаты, необходимо строго соблюдать заданную температуру.
Принцип метода. Для определения осахаривающей способности, характеризующей активность β– амилазы, проводят гидролиз растворимого крахмала исследуемым ферментным препаратом при рН 5,6 и различной температуре. Количество образовавшегося сахара мальтозы определяют методом Бертрана.
Метод Бертрана основан на способности свободной альдегидной или кетонной группы молекулы сахара взаимодействовать со щелочным раствором окисной меди (II) (реактивом Фелинга) и восстанавливать ее до оксида меди (I), выпадающего в виде осадка красного цвета.
По количеству образовавшегося оксида меди (I) судят о содержании сахара в испытуемом растворе.
Реактив Фелинга представляет собой смесь разных объемов 4 %–ной сернокис лой меди и щелочного раствора сегнетовой соли. При смешивании сернокислой меди со щелочью выпадает осадок гидрата окиси меди (II). Сегнетова соль препятствует выпадению осадка, образуя комплексное растворимое соединение.
21
|
|
|
|
Cu(OH)2 + 2 NaOH |
|
|
|
Cu(OH)2 + Na2SO4 |
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|||||||||||
|
|
O |
|
O |
|
|
|
|
|
O |
|
|||||||
HO |
|
|
|
|
|
|
|
|
|
|
OH |
O |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
CH |
|
|
ONa |
NaO |
|
CH |
Cu |
|
CH |
|
|
ONa + 2H O |
|||||
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
CH |
|
|
OK |
KO |
|
CH |
|
|
CH |
|
|
OK |
|
|||||
HO |
|
|
|
|
|
|
|
|
|
|
O |
HO |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
O |
|
|||||||||
|
|
|
O |
|
|
|
|
|
|
|||||||||
В щелочной среде циклическая (полуацетальная) форма сахара полностью переходит в ациклическую и на месте свободного гликозидного гидроксила образу-
CH2OH |
|
CH2OH |
CH2OH |
|
|||
H |
O H |
H |
OHH |
O H |
H |
O H |
H |
OHH |
H |
|
H |
OHH |
H |
|
|
HO |
|
O |
|
OH |
HO |
|
O |
H |
OH |
|
H |
OH |
H |
OH |
|
|
|
|
|
||||
|
|
|
|
|
|
||
ется альдегидная (у альдоз) или кетонная (у кетоз) группа.
|
|
CH2OH |
|
|
|
|
|
OH |
O |
|
|
|
||
|
H |
|||
|
C |
|||
OH |
|
|
H |
|
|
|
H |
||
|
|
|
|
|
HOH
|
|
|
|
|
O |
|
|
|
|
O |
|
|
|
|
|
|
|
|
O |
|||||||
|
|
O |
|
|
|
|
|
OH |
|
O |
|
CH |
|
|
ONa + 2H2O |
|
|
|
|
O |
HO |
|
CH |
|
|
ONa |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
NaO |
|
|
|
CH |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
R |
|
C |
+ |
|
|
|
Cu |
|
|
|
|
|
R |
|
C |
+ |
2 |
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OK |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
H |
KO |
|
|
|
CH O |
|
HO |
|
CH |
|
|
OK |
|
|
|
|
OH |
HO |
|
CH |
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
O |
|
|
|
|
O |
|
|
|
|
+ Cu2O |
|
|
O |
||||||||
Так, например, превращение циклической формы мальтозы в ациклическую протекает следующим образом:
Взаимодействие сахаров с реактивом Фелинга протекает по уравнению:
Количество образовавшегося осадка оксида меди (I) строго соответствует количеству сахара, находящегося в растворе, и определяется объемным методом. Для этого предварительно отмытый от избытка реактива Фелинга осадок оксида меди (I) обрабатывают раствором железоаммиачных квасцов. При этом медь (I) переходит в медь (II), а эквивалентное количество железа (III) восстанавливается до железа (II).
Сu+ – е ¾® Cu++ Fе+++ + е ¾® Fе++
Количество восстановленного железа, эквивалентное количеству оксида меди (I), определяют титрованием раствором перманганата. Весь процесс сводится к следующим реакциям:
Сu2О + Fе2(SО4)3 + Н2SО4––> 2 СuSО4 + 2 FеSО4 + Н2О;
22
10 FеSО4+2 КМnО4 + 8 Н2SО4––> 2Fе2(SО4)3+ К2SО4 +2 МnSО4 + 8 Н2О
Титр перманганата устанавливают по меди, что дает возможность сразу пересчитать объем пошедшего на титрование перманганата на эквивалентное количество миллиграммов оксида меди (I).
Ход работы. В колбу вносят 10 мл 2 % –ного крахмального клейстера и 2 мл фосфатного буфера рН 5,6. Содержимое колбы доводят до заданной температуры (нагревают или охлаждают), после чего вносят 1 мл ферментной вытяжки, предварительно термостатированной при той же температуре, и замечают время. Гидролиз проводят 30 минут, в течение которых строго следят за поддержанием заданной температуры. Через 30 мин. в колбу вносят 3 мл 6 %– ного раствора сульфата меди для прекращения действия фермента, содержимое колбы охлаждают и доводят водой до 20 мл. Количество образовавшейся мальтозы определяют методом Бертрана. Для этого в коническую колбу пипеткой отбирают 10 мл полученного гидролизата, добавляют 10 мл воды и 40 мл реактива Фелинга (смесь 20 мл Фелинга 1 и 20 мл Фелинга II).
Испытуемый раствор с жидкостью Фелинга нагревают до кипения и кипятят 3 минуты. Кипение должно идти спокойно, жидкость в колбе должна оставаться синей, что указывает на избыток реактива Фелинга, а, следовательно, и на полноту окисления.
Сняв раствор с огня, дают некоторое время для осаждения оксида меди (I), затем жидкость декантируют через асбестовый фильтр в колбу Бунзена, соединенную с отсасывающим насосом. Оксид меди (I) должен остаться в колбе, но часть его может перейти на фильтр. Осадок в колбе 3–4 раза промывают небольшими порциями горячей воды, декантируя через тот же асбестовый фильтр. Осадок в колбе и на фильтре, во избежание его окисления кислородом воздуха, должен все время находиться под жидкостью.
После отмывания осадка Сu2О от щелочи колбу Бунзена отключают от насоса, содержимое ее выливают. В чисто промытую колбу Бунзена вставляют тот же асбестовый фильтр. Промытый осадок оксида меди (I) в колбочке растворяют 15–20 мл железоаммиачных квасцов и количественно переносят на асбестовый фильтр. Для лучшего растворения осадка на фильтре верхний слой асбеста слегка разрыхляют стеклянной палочкой.
Когда осадок растворится, соединяют колбу Бунзена с насосом и отсасывают жидкость с фильтра. Колбочку, где был осадок, тщательно ополаскивают несколько раз водой, и промывные воды отсасывают в колбу Бунзена. Зеленоватый раствор в колбе Бунзена, содержащий ионы железа (II) , титруют раствором КМnО4 до слаборозовой окраски, не исчезающей в течение одной минуты. Израсходованное на титрование количество мл 0,1 н. раствора перманганата (с учетом поправки на титр) умножают на его титр по меди (6,36 мг Си2О) и по таблице (см. Приложение 2, таб.3) определяют количество мальтозы.