

56. Клеточный цикл и деление клеток – митоз и мейоз.
Мейоз — это деление в зоне созревания половых клеток, сопровождающееся уменьшением числа хромосом вдвое. Он состоит из двух последовательно идущих делений, имеющих те же фазы, что и митоз. Однако, как показано в таблице «Сравнение митоза и мейоза», продолжительность отдельных фаз и происходящие в них процессы значительно отличаются от процессов, происходящих при митозе.
Эти отличия в основном состоят в следующем.
В мейозе профаза I более продолжительна. В ней происходит конъюгация(соединение гомологичных хромосом) и обмен генетической информацией. В анафазе Iцентромеры, скрепляющие хроматиды, не делятся, а к полюсам отходит одна из гомологмейоза Митоз и его фазы митоза и ичных хромосом. Интерфаза перед вторым делением очень короткая, в ней ДНК не синтезируется. Клетки (галиты), образующиеся в результате двух мейотических делений, содержат гаплоидный (одинарный) набор хромосом. Диплоидность восстанавливается при слиянии двух клеток — материнской и отцовской. Оплодотворенную яйцеклетку называютзиготой.
Митоз, или непрямое деление, наиболее широко распространен в природе. Митоз лежит в основе деления всех неполовых клеток (эпителиальных, мышечных, нервных, костных и др.). Митоз состоит из четырех последовательных фаз (см. далее таблицу). Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними. Период жизни клетки между двумя митозами называют интерфазой. Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков, удваивается каждая хромосома, образуя две сестринские хроматиды, скрепленные общей центромерой, увеличивается число основных органоидов цитоплазмы.
В профазе спиралируются и вследствие этого утолщаются хромосомы, состоящие из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредоточиваются по всей клетке, центриоли отходят к полюсам и образуют веретено деления. В метафазе происходит дальнейшая спирализация хромосом. В эту фазу они наиболее хорошо видны. Их центромеры располагаются по экватору. К ним прикрепляются нити веретена деления.
В анафазе центромеры делятся, сестринские хроматиды отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки.
В телофазе цитоплазма делится, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. В животных клетках цитоплазма перешнуровывается, в растительных — в центре материнской клетки образуется перегородка. Так из одной исходной клетки (материнской) образуются две новые дочерние.
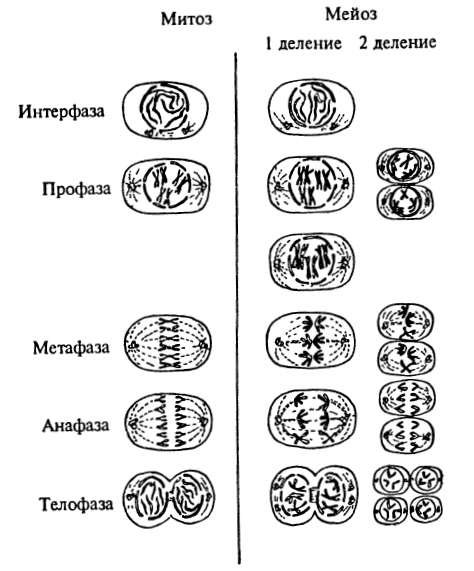
Мейоз и митоз
Таблица - Сравнение митоза и мейоза
|
Фаза |
Митоз |
Мейоз | |
|
1 деление |
2 деление | ||
|
Интерфаза |
Набор хромосом 2n Идет интенсивный синтез белков, АТФ и других органических веществ Удваиваются хромосомы, каждая оказывается состоящей из двух сестринских хроматид, скрепленных общей центромерой. |
Набор хромосом 2n Наблюдаются те же процессы, что и в митозе, но более продолжительна, особенно при образовании яйцеклеток. |
Набор хромосом гаплоидный (n). Синтез органических веществ отсутствует. |
|
Профаза |
Непродолжительна, происходит спирализация хромосом, исчезают ядерная оболочка, ядрышко, образуется веретено деления |
Более длительна. В начале фазы те же процессы, что и в митозе. Кроме того, происходит конъюгация хромосом, при которой гомологичные хромосомы сближаются по всей длине и скручиваются. При этом может происходить обмен генетической информацией (перекрест хромосом) —кроссинговер. Затем хромосомы расходятся. |
Короткая; те же процессы, что и в митозе, но при nхромосом. |
|
Метафаза |
Происходит дальнейшая спирализация хромосом, их центромеры располагаются по экватору. |
Происходят процессы, аналогичные тем, что и в митозе. |
Происходит то же, что и в митозе, но при nхромосом. |
|
Анафаза |
Центромеры, скрепляющие сестринские хроматиды, делятся, каждая из них становится новой хромосомой и отходит к противоположным полюсам. |
Центромеры не делятся. К противоположным полюсам отходит одна из гомологичных хромосом, состоящая из двух хроматид, скрепленных общей центромерой. |
Происходит то же, что и в митозе, но при nхромосом. |
|
Телофаза |
Делится цитоплазма, образуются две дочерние клетки, каждая с диплоидным набором хромосом. Исчезает веретено деления, формируются ядрышки. |
Длится недолго Гомологичные хромосомы попадают в разные клетки с гаплоидным набором хромосом. Цитоплазма делится не всегда. |
Делится цитоплазма. После двух мейотических делений образуется 4 клетки с гаплоидным набором хромосом. |
Клеточный цикл — это период существования клетки от момента её образования путем деления материнской клетки до собственного деления.
Длительность клеточного цикла эукариот
Длительность клеточного цикла у разных клеток варьируется. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12—36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих, земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10—24 ч.
Фазы клеточного цикла эукариот
Клеточный цикл эукариот состоит из двух периодов:
Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Периода клеточного деления, называемый «фаза М» (от слова mitosis — митоз).
Интерфаза состоит из нескольких периодов:
G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов;
S-фазы (от англ. synthesis — синтез), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть).
G2-фазы, во время которой идет подготовка к митозу.
У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
Период клеточного деления (фаза М) включает две стадии:
-кариокинез (деление клеточного ядра);
-цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий.
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатахсветовой и электронной микроскопии фиксированных и окрашенных клеток.
Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, какциклин-зависимые киназы и циклины. Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на нихфакторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своимирецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса.Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклинаB в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Контрольные точки клеточного цикла
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку, то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт. Существует как минимум четыре контрольных точки клеточного цикла: точка в G1, где проверяется интактность ДНК, перед вхождением в S-фазу, сверочная точка в S-фазе, в которой проверяется правильность репликации ДНК, сверочная точка в G2, в которой проверяются повреждения, пропущенные при прохождении предыдущих сверочных точек, либо полученные на последующих стадиях клеточного цикла. В G2 фазе детектируется полнота репликации ДНК, и клетки, в которых ДНК недореплицирована, не входят в митоз. В контрольной точке сборки веретена деления проверяется, все ли кинетохоры прикреплены к микротрубочкам.
Нарушения клеточного цикла и образование опухолей
Увеличение синтеза белка p53 ведет к индукции синтеза белка p21 — ингибитора клеточного цикла
Нарушение нормальной регуляции клеточного цикла является причиной появления большинства твердых опухолей. В клеточном цикле, как уже говорилось, прохождение контрольных пунктов его возможно только в случае нормального завершения предыдущих этапов и отсутствия поломок. Для опухолевых клеток характерны изменения компонентов сверочных точек клеточного цикла. При инактивации сверочных точек клеточного цикла наблюдается дисфункция некоторых опухолевых супрессоров и протоонкогенов, в частности p53, pRb, Myc иRas. Белок p53 является одним из факторов транскрипции, который инициирует синтез белка p21, являющегося ингибитором комплекса CDK-циклин, что приводит к остановке клеточного цикла в G1 и G2 периоде. Таким образом клетка, у которой повреждена ДНК, не вступает в S-фазу. При мутациях, приводящих к потере генов белка p53, или при их изменениях, блокады клеточного цикла не происходит, клетки вступают в митоз, что приводит к появлению мутантных клеток, большая часть из которых нежизнеспособна, другая — дает начало злокачественным клеткам.
Деление клеток
|
|
|
Деление бактериальной клетки |
Все клетки появляются путём деления родительских клеток. Большинству клеток свойственен клеточный цикл, состоящий из двух основных стадий: интерфазы и митоза.
Интерфаза состоит из трех этапов. В течение 4–8 часов после рождения клетка увеличивает свою массу. Некоторые клетки (например, нервные клетки мозга) навсегда остаются в этой стадии, у других же в течение 6–9 часов удваивается хромосомная ДНК. Когда масса клетки увеличивается в два раза, начинается митоз.
|
|
В типичной животной клетке митоз происходит следующим образом. В профазу центриоли удваиваются, две образовавшиеся центриоли начинают расходиться к разным полюсам клетки. Ядерная мембрана разрушается. Специальные микротрубочки выстраиваются от одной центриоли к другой, образуя веретено деления. Хромосомы разъединяются, но всё ещё остаются попарно сцепленными.
Следующая после профазы стадия называется метафазой. Хромосомы, влекомые нитями веретена, выстраиваются в экваториальной плоскости клетки. Центромеры, скреплявшие хромосомы, делятся, после чего дочерние хромосомы полностью разъединяются.
|
|
|
|
В стадии анафазы хромосомы перемещаются к полюсам клетки. Когда хромосомы достигают полюсов, начинается телофаза. Клетка делится надвое в экваториальной плоскости, нити веретена разрушаются, вокруг хромосом формируются ядерные мембраны. Каждая дочерняя клетка получает собственный набор хромосом и возвращается в стадию интерфазы. Весь процесс занимает около часа.
Процесс митоза может варьировать в зависимости от типа клетки. В растительной клетке отсутствуют центриоли, хотя веретено деления образуется. В грибных клетках митоз происходит внутри ядра, ядерная мембрана не распадается.
|
|
|
|
Наличие хромосом не является необходимым условием деления клетки. С другой стороны, один или несколько митозов могут останавливаться на стадии телофазы, в результате чего возникают многоядерные клетки (например, у некоторых водорослей).
Размножение при помощи митоза называют бесполым или вегетативным, а также клонированием. При митозе генетический материал родительских и дочерних клеток идентичен.
Мейоз, в отличие от митоза, является важным элементом полового размножения. При мейозе образуются клетки, содержащие лишь один набор хромосом, что делает возможным последующее слияние половых клеток (гамет) двух родителей. По сути, мейоз является разновидностью митоза. Он включает два последовательных деления клетки, однако хромосомы удваиваются только в первом из этих делений. Биологическая сущность мейоза заключается в уменьшении числа хромосом в два раза и образовании гаплоидных гамет (то есть гамет, имеющих по одному набору хромосом).
В результате мейотического деления у животных образуются четыре гаметы. Если мужские половые клетки имеют примерно одинаковые размеры, то при образовании яйцеклеток распределение цитоплазмы происходит очень неравномерно: одна клетка остаётся крупной, а три остальных настолько малы, что их почти целиком занимает ядро. Эти мелкие клетки служат лишь для размещения избыточного генетического материала.
Мужские и женские гаметы сливаются, образуя зиготу. Хромосомные наборы при этом объединяются (этот процесс называется сингамией), в результате чего в зиготе восстанавливается удвоенный набор хромосом – по одному от каждого из родителей. Случайное расхождение хромосом и обмен генетическим материалом между гомологичными хромосомами приводят к возникновению новых комбинаций генов, повышая генетическое разнообразие. Образовавшаяся зигота развивается в самостоятельный организм.
В последнее время проводились эксперименты по искусственному слиянию клеток одного или разных видов. Наружные поверхности клеток склеивались вместе, а мембрана между ними разрушалась. Таким образом удалось получить гибридные клетки мыши и цыплёнка, человека и мыши. Однако при последующих делениях клетки теряли большинство хромосом одного из видов.
В других экспериментах клетка разделялась на компоненты, например, ядро, цитоплазму и мембрану. После этого компоненты различных клеток снова соединяли вместе, и в результате получалась живая клетка, состоящая из компонентов клеток разных видов. В принципе, опыты по сборке искусственных клеток могут оказаться первым шагом на пути к созданию новых форм жизни.