

Вопрос1. Биосинтез жирных кислот
Наряду с расщеплением жирных кислот в результате их окисления, в клетках функционирует процесс синтеза жирных кислот. Путь синтеза жирных кислот не является обратным пути их расщепления. Он представляет собой новую последовательность реакций.
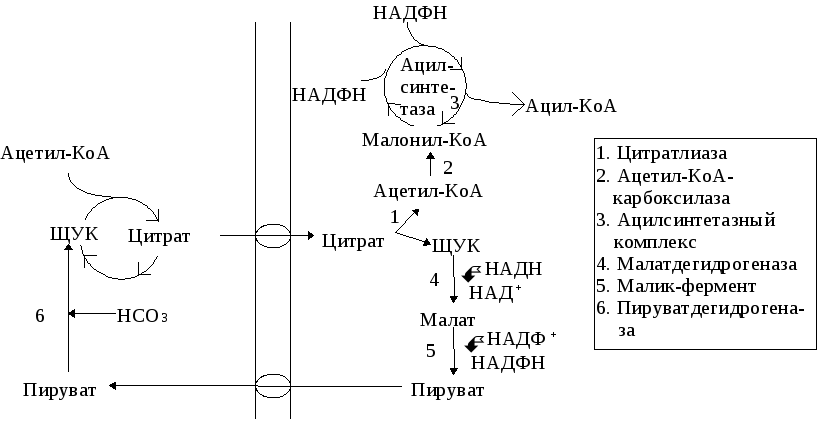
Схема перемещения ацетил-КоА из митохондрий в цитозоль
Основными особенностями пути биосинтеза жирных кислот являются следующие:
1. Синтез происходит в цитоплазме в отличие от распада, который протекает в матриксе митохондрий.
2. Промежуточные продукты синтеза жирных кислот ковалентно связаны с ацилпереносящим белком (АПБ), тогда как промежуточные продукты окисления жирных кислот связаны с коферментом А.
3. Большинство ферментов синтеза жирных кислот организованы в мультиферментный комплекс, называемый синтетазой жирных кислот.
4. Удлинение цепи синтезируемой жирной кислоты происходит путем последовательного присоединения двухуглеродных фрагментов. Поставщиком этих двухуглеродных фрагментов служит малонил-АПБ.
5. В качестве восстановителя при синтезе жирных кислот выступает НАДФН.
6. На этапе образования пальмитата (С16) удлинение остатка жирных кислот с участием мультиферментного комплекса синтетазы жирных кислот останавливается. Дальнейшее удлинение, как и введение двойных связей, происходят под влиянием других ферментных систем.
Синтез жирных кислот de novo происходит при наличии достаточного количества глюкозы, когда ацетил-КоА образуется больше, чем требуется для покрытия энергетических нужд. В этих условиях избыток моносахаридов, образовавшийся в результате переваривания углеводов, может запасаться не в форме водорастворимого гликогена, а в виде гидрофобных ТАГ.
Субстраты синтеза жирных кислот. Предшественником является ацетил-КоА, процесс протекает в цитозоле. Ацетил-КоА образуется из пирувата под действием митохондриального пируватдегидрогеназного комплекса. Но внутренняя митохондриальная мембрана непроницаема для ацетил-КоА. В митохондриях фермент цитратсинтаза катализирует реакцию образования цитрата из ацетил-КоА и ЩУК. Цитрат выходит из митохондрий в цитоплазму. В цитозоле фермент АТФ-цитратлиаза расщепляет цитрат до ацетил-КоА и ЩУК (см рис). Этот ацетил-КоА и принимает участие в синтезе жирных кислот.
Ферменты синтеза жирных кислот. В биосинтезе насыщенных жирных кислот участвуют два ферментных комплекса: ацетил-КоА карбоксилаза и ацилсинтетаза. В случае синтеза пальмитиновой кислоты необходимы 8 молекул ацетил-КоА, 14 НАДФН (в качестве восстановительных эквивалентов) и 7 АТФ. Первая молекула ацетил-КоА действует как включатель ацилсинтетазы. Остальные двухуглеродные фрагменты вовлекаются в процесс после того, как молекула ацетил-КоА превращается в малонил-КоА под действием ацетил-КоА карбоксилазы.
Ацетил-КоА карбоксилаза, содержащая в качестве простетической группы биотин (витамин Н), катализирует ключевую реакцию в синтезе жирных кислот. Эта реакция выглядит следующим образом:

|
|
|
|
Механизм действия ацетил КоА карбоксилазы | |
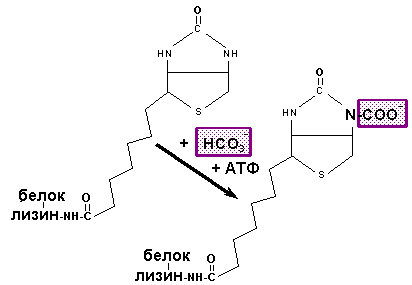

Имеется несколько механизмов регуляции активности ацетил-КоА карбоксилазы. Есть мономерная (неактивная) и полимерная (активная) формы фермента. Цитрат активирует фермент за счет увеличения образования полимерной формы, содержащей около 20 мономерных субъединиц. Ацетил-КоА карбоксилаза подвергается обратимому фосфорилированию/дефосфорилированию; цАМФ-зависимая протеинкиназа ингибирует ферментативную активность, а фосфатаза - активирует. На этом основана гормональная регуляция активности ацетил-КоА карбоксилазы. Повышенные концентрации малонил-КоА и ацил-КоА аллостерически ингибируют активность этого фермента.
Синтетаза жирных кислот - это мультиферментный комплекс, включающий в себя 7 ферментов и белок, содержащий в своем составе 4-фосфопантотеиновую простетическую группу. Суммарная молекулярная масса этого комплекса, находящегося в цитоплазме клеток, составляет около 400 тыс.Да. Центральное место в этой системе занимает ацилпереносящий белок, с которым ковалентно связываются промежуточные продукты биосинтеза жирных кислот.

Роль простетической группы в АПБ выполняет фосфопантотеин, который также входит в состав кофермента А. Функция АПБ в биосинтезе жирных кислот аналогична функции кофермента А в окислении жирных кислот. В процессе построения цепи жирной кислоты промежуточные продукты образуют эфирные связи с АПБ. Белок является своеобразной сердцевиной комплекса и функционирует как переносчик промежуточных продуктов в растущей цепи ацила. Синтез инициируется праймером (первой молекулой ацетил-КоА), с которой взаимодействует малонил-КоА, при этом выделяется СО2 и образуется ацетоацетильный фрагмент. Последний восстанавливается при участии НАДФН+Н+ и после отщепления воды превращается в еноильный остаток. Этот остаток вновь восстанавливается при участии НАДФН+Н+ в остаток масляной кислоты.
Началом последующего цикла, который включает в себя ту же последовательность реакций, является конденсация бутирил-АПБ с следующей молекулой малонил-АПБ. Добавление каждого двухуглеродного фрагмента в растущую ацильную цепь сопровождается циклом реакций конденсации, восстановления (используется НАДФН), дегидратации и восстановления (используется вторая молекула НАДФН). За 7 таких циклов образуется насыщенная жирная кислота с 16 углеродными атомами. В общем виде это выглядит следующим образом:
ацетил~КоА + 7 малонил~КоА + 14 НАДФН + 14 Н+ > пальмитат + 7 СО2 + 14 НАДФ + 8 КоА-SH + 6 Н2О
Потребность в НАДФН для реакций восстановления в процессе синтеза жирных кислот. НАДФН+Н+, который необходим для восстановительных реакций, протекающих в ходе биосинтеза жирных кислот, в различных клетках поступает из двух различных источников. В печени НАДФН+Н+ образуется, главным образом, в реакциях пентозофосфатного пути. В клетках жировой ткани НАДФН образуется преимущественно за счет функционирования челночного механизма, обеспечивающего перенос остатка уксусной кислоты из митохондрий в цитоплазму. Последний протекает с участием фермента малатдегидрогеназы (кофермент НАДН) и малик фермента (в реакции восстановления НАДФ) (схему см. выше).
Как уже отмечалось, при участии синтетазы жирных кислот идет образование ацилов, содержащих максимально 16 углеродных атомов. Дальнейшее удлинение цепей жирных кислот происходит при помощи других ферментных систем путем присоединения ацетильных групп; этот процесс протекает в эндоплазматическом ретикулуме и митохондриях. Скорость биосинтеза жирных кислот определяется, главным образом, скоростью ацетил-КоА-карбоксилазной реакции.