

2.3-дифосфоглицерат сдвигает кривую насыщения гемоглобина кислородом вправо
Эритроцит обладает системой механизмов направленных на поддержание обеспечения кислородом тканей при различных неблагоприятных
внешних условиях. Так при пребывании на высоте в 4500 м наблюдается выраженное повышение
концентрации 2,3- дифосфоглицерата, которое нормализуется через 52 часа после возвращения человека к обычным условиям. Одновременно с
повышением 2,3 дифосфоглицерата повышается Р50, что соответствует сдвигу кривой насыщения вправо. Организм реагирует таким же
компенсаторным механизмом на изменения количества эритроцитов в циркуляции.
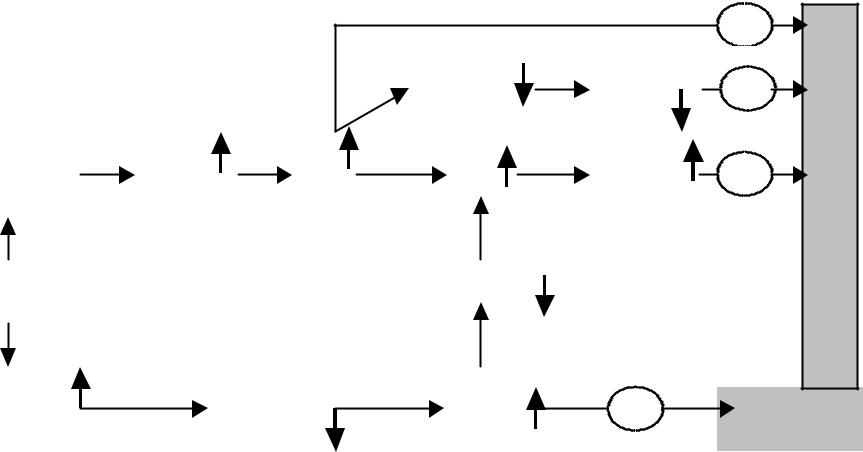
Гипоксия ведет к повышению уровня 2,3 дифосфоглицерата в эритроцитах. Изменение рН
в эритроцитах играет ключевую роль в регуляции уровня 2,3-дифосфоглицерата при гипоксии(недостатке кислорода в тканях), алкалозе (повышение рН во внеклеточном пространстве) или ацидозе (падение рН во внеклеточном пространстве).
При гипоксии недостаток кислорода ведет к гипервентиляции с повышенным выделением СО2, что сопровождается повышением значения рН крови и эритроцитов (алкалоз). В эритроцитах алкалоз поддерживается также повышенным связыванием протонов дезоксигемоглобином, количество которого увеличивается при гипоксии
|
|
|
|
|
- |
|
|
|
|
2,3-ДФГ |
Распад 2,3- |
+ |
|
|
|
|
фосфатаза |
ДФГ |
||
|
|
|
|
|||
Гипер |
рН |
рН |
ФФК |
1,3-ДФГ |
+ |
|
вентиляция |
крови |
эритр |
||||
|
|
|
Гипоксия |
АТФ/АДФ |
Дезокси ге- |
Свободный |
2,3-ДФГ |
+ |
2,3-ДФГ |
|
моглобин |
2,3-ДФГ |
мутаза |
|||
|
|
Механизм повышения уровня 2,3-дифосфоглицерата, индуцируемого гипоксией. ДФГ- дифосфоглицерат, ФФК-фосфофруктокиназа
Защелачивание внутри эритроцитов ведет к повышению активности ФФК и усилению образования 1,3-ДФГ, а следовательно, и 2,3- ДФГ .Одновременно, тормозится активность фосфатазы 2,3-ДФГ, что является дополнительным источником повышения его концентрации в эритроцитах. Однако, повышение уровня 2,3-ДФГК ведет к увеличению уровня протонов в эритроцитах и противодействует вызванному гипоксией повышению рН внутри эритроцитов и изменениям, связанным с повышением рН, что составляет основу принципа обратной связи, согласно которому 2,3-ДФГ регулирует свой уровень в эритроцитах
Эритроциты постоянно разрушаются, заменяясь новыми. Суточный распад- 20 мл Э, т.е. 0.85% всей массы Э.
Каждый час у взрослого образуется 6×109 степени кр .кр клеток. Период жизни Э 110-120 дней. У 80 дневного Э концентрация МетHb в 4-5 раз больше, чем у зрелого. Изменение структуры мембраны меняет форму и приводит к гемолизу.
Распад Э начинается в периферической крови и заканчивается в клетках РЭС, где образуется непрямой Билирубин. Клетками РЭС и кровью билирубин переносится к печени. Билирубин представляет собой неполярное, нерастворимое в воде соединение. Оно связывается с альбумином и не проходит через почечный барьер. Под действием микросомальной гем-оксигеназы происходит окисление гема.
Гем +О2 --- биливердин + СО2 + Fe ++
Коферментом микросомальной гем- оксигеназы является NADF*H. Микросомальная гем- оксигеназа имеет смешанную функцию. Для нее необходимы- О2, NADF*H и цитохром С-редуктаза.
Гем +О2 --- биливердин + СО2 + Fe ++
Гем переходит в вердоглобин далее в биливердин и билирубин.
Образующиеся катионы Fe ++ захватывается трансферрином и направляется в депо, или в костный мозг, на биосинтез Hb или других гем содержащих протеидов.
Глобин подвергается гидролизу на аминокислоты и также используется при других биосинтезах.
Образующийся при распаде гемоглобина билирубин поступает в
кровь и связывается альбуминами плазмы - это непрямой билирубином. В отличие от прямого билирубина,
непрямой не даёт цветную реакцию с диазореактивом Эрлиха пока белки плазмы крови не осаждены спиртом.
Непрямой билирубин называется также свободным, поскольку его связь с
альбуминами плазмы не химическая (не ковалентная), а абсорбционная.
Клетками РЭС и кровью билирубин переносится к печени. Билирубин переносится в соотношении --1моль А на 2 моля Б.На клеточных мембранах печени, Билирубин частично освобождается от Альбумина и переносится в гепатоциты. Билирубин это неполярное, нерастворимое в воде соединение. Связанное с А , оно не проходит через почечный барьер. Под действием микросомальной гем- оксигеназы происходит окисление гема.
Б поступает в кишечник через желчные протоки и под действием кишечных бактерий и анаэробных дегидрогеназ- восстанавливается в уробилиноген и стеркобилиноген -основные пигменты мочи и кала.
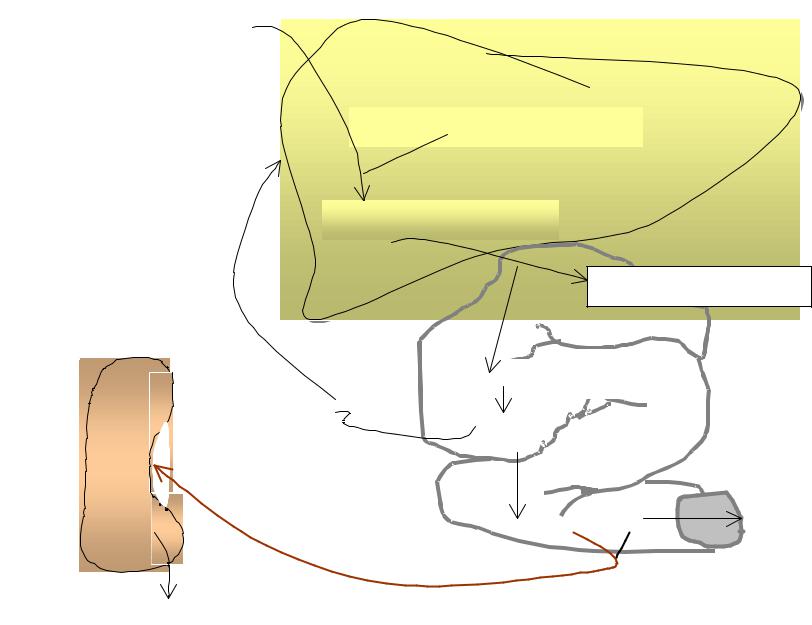
Непрямой билирубин |
2УДФ-глюкуроновая к-та |
Прямой билирубин |
2УДФ-глюкурон.к-та |
Мезобилирубин |
Уробилин |
Стеркобилин |
Стеркобилиноген |