

Между обменами глюкозы и этанола в печени
2.2. Роль печени в метаболизме липидов
1.В печени образуется жёлчь. Компонентами жёлчи, кроме воды, явля-ются соли жёлчных кислот, холестерин, фосфолипиды, билирубин-глюкурониды, муцин, немного белков (альбумины, 5’-нуклеотидаза, щелочная фосфатаза, ГГТП, иммуноглобулин А), различных ионов (Na+, Cl-, HCO3-, Cu++, Fe+++, Se++ и др.), микроколичества липовитаминов, стероидных гормонов и т.д.
Основные функции жёлчи: а) пищеварительная – соли жёлчных кислот эмульгируют липиды в тонком кишечнике, активируют липазы, создают оптимум рН для их действия, служат основными компонентами холеиновых комплексов – специфических частиц, в составе которых происходит всасывание ВЖК, ХС, липовитаминов; б) выделительная – попадая в просвет кишечника, некоторые из составных компонентов жёлчи выделяются с калом из организма; правда, часть из них преобразуется микрофлорой кишечника: билирубин в уробилин (стеркобилин).
ВЖК-альбумины ЛПЛ


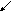 плазмы
плазмы


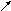
 ЛП
плазмы ЛХАТ
ЛП
плазмы ЛХАТ
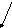 крови
крови






 Глицерофосфатиды
ВЖК
печени
ТАГ ЛП
Глицерофосфатиды
ВЖК
печени
ТАГ ЛП
печени плазмы

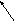 Фосфолипиды
крови
Фосфолипиды
крови

 Ацил-КоА
Ацил-КоА
 ВЖК
ВЖК
ТАГ печени
 Ацетил-КоА
Холестерин
Ацетил-КоА
Холестерин


ЦТК
 Соли
Жёлчь
Соли
Жёлчь
жёлчных
кислот
 Кетоновые
Кетоновые
Кетоновые
Кетоновые
тела тела плазмы крови
Схема 3. Судьба высших жирных кислот в печени (ключевые блоки)
2.Центральное место обмена ВЖК (схема 3). Сюда они поступают из кишечника, жировых депо в составе альбуминов плазмы крови. Здесь часть жирных кислот синтезируется из ацетил-КоА, получившегося путём многостадийного преобразования глюкозо-6-фосфата с участием восстановленного НАДФ. Мало того, в печени есть специальная ферментативная система, которая может внедрить одну двойную связь в середину цепи стеариновой кислоты, превращая её в олеиновую; а также удлинять и дегидрировать линолевую кислоту с образованием арахидоновой.
3.В митохондриях печени синтезируются из ацетил-КоА (при дефиците оксалоацетата) β-гидроксимасляная, ацетоуксусная кислоты, которые,
Глюкозо-6-фосфат Ацил КоА
 ВЖК
ВЖК

 ПВК
ПВК
ОА Ацетил КоА







 ЦТК
ЦТК
Цитрат

 β-гидрокси-,
β-гидрокси-,
β-метил-глутарил-КоА


 ацетоацетат
Общий
кровоток
ацетоацетат
Общий
кровоток
 НАДН+Н+
НАДН+Н+
Кетоновые тела
НАД+
β-гидрокси-бутират
Схема 4. Синтез кетоновых тел в митохондриях гепатоцитов
легко преодолевая мембраны, оказываются в общем кровотоке (клинический термин – кетоновые тела) (схемы 3,4).
4.В печени из метаболитов гликолиза дигидроксиацетонфосфата или реже глицеральдегид-3-фосфата – образуется глицерол-1фосфат, последний необходим в синтезе фосфатидата – соединения, из которого в зависимости от обстоятельств могут синтезироваться нейтральные жиры или глицерофосфатиды.
Для генеза данного ключевого предшественника может использоваться и глицерин, поступивший в гепатоциты с кровью. Только он должен предварительно фосфорилироваться. Реакция катализируется глицеролкиназой. В отличие от печени в липоцитах активность данного фермента не регистрируется. Последнее свидетельствует о том, что источником глицеролфосфата в жировой ткани служит только продукт преобразования глюкозы. Иными словами, нейтральные жиры в липоцитах получаются, в основном, из этого моносахарида.
5.Если в плазме крови происходит хотя бы временное увеличение уровня ВЖК, они, поступая в гепатоциты, взаимодействуют с фосфатидной кислотой с образованием нейтральных жиров (схема 3).
6.При достаточном количестве в гепатоцитах холина, этаноламина, серина, инозитола, последние могут включаться в фосфатидную кислоту, что заканчивается синтезом соответствующих глицерофосфатидов (липотропных веществ).
7.В печени образуется часть апопротеинов – белковых компонентов ЛП, в первую очередь, ЛПОНП, ЛПНП.
8.В гепатоцитах синтезируется ЛПОНП – основные транспортные формы эндогенных нейтральных жиров. Под влиянием липопротеинлипазы, локализующейся обычно на стенках капилляров и высвобождающейся в кровяное русло при участии гепарина, происходит постадийный гидролиз жиров ЛПОНП:
ЛПЛ ЛПЛ ЛПЛ




 ТАГ ДАГ
МАГ Глицерол
ТАГ ДАГ
МАГ Глицерол
ВЖК ВЖК ВЖК
Часть высвободившихся ВЖК связывается сывороточным альбумином, другая же, преодолевая цитолеммы, попадает в ткани, а ЛПОНП, теряя их, преобразуются постепенно в ЛПНП (схема 3).
9.Печень, в меньшей степени кишечник, – наиболее активные продуценты холестерина, который поступает в общий кровоток в составе ЛПНП или ЛПОНП (реже). Происхождение последних имеет альтернативу (см. выше). Именно ЛПНП распознаются рецепторами клеток периферических тканей и поглощаются ими путём эндоцитоза, снабжая их остатками ТАГ-ов и холестерином (схема 3).
10.В гепатоцитах образуются и ЛПВП - дископодобные структуры, включающие апобелки, фосфолипиды и относительно малые количества ХС и нейтральных жиров. В периферических тканях они обогащаются холестерином, причём обычно гидроксил этого спирта экспонирован в водное окружение ЛПВП и доступен для этерификации. Чтобы амфипатичный холестерин стал полностью гидрофобным и был способен мигрировать с периферии ЛП к центру, тем самым снизив атерогенность частицы, требуется работа ЛХАТ.
11.Лецитин-холестерин-ацилтрансфераза (ЛХАТ) – фермент, синтезируемый печенью, но катализирующий перенос ВЖК от фосфатидилхолина (старое название – лецитин) на холестерин в ЛПНП плазмы крови (схема 3).
12.Таким образом, печень служит главным органом в метаболизме холестерина. В ней он синтезируется, этерифицируется, включается в соответствующий ЛП, отправляющийся к периферическим тканям, сюда же холестерин возвращается, чтобы экскретироваться в свободном виде в составе жёлчи или же предварительно преобразовавшись в жёлчные кислоты.