

2.1. Участие печени в углеводном обмене
1.Этот орган служит «диспетчером» для всосавшихся и доставленных по воротной вене простых углеводов. В зависимости от природы, их содержания в общем кровотоке, потребностей организма дальнейшая судьба моносахаридов различна.
2.Часть их отправится в печёночную вену, чтобы поддержать гомеостаз, в первую очередь, глюкозы крови и обеспечить нужды органов. Часть печень использует для собственных нужд. Оставшиеся моносахариды будут депонироваться или в виде гликогена (его накопление ограничено из-за солидной молекулярной массы), или же в виде нейтральных жиров.
В гепатоцитах интенсивно протекает метаболизм глюкозы. Поступившие с пищей моносахариды (галактоза, манноза, фруктоза и др.) только в печени с помощью специфических ферментных систем преобразуются в глюкозо-6-фосфат (лишь в такой форме глюкоза используется клетками). Всего в двух органах (печени, в меньшей степени, в почках) регистрируется активность энзимов глюконеогенеза – процесса, осуществляющего синтез глюкозы из продуктов неуглеводного происхождения (аминокислот, пирувата, лактата, глицерина и др.). Печень – единственный орган, где лактатдегидрогеназа (ЛДГ) работает в прямой реакции:
НАД+НАДН + Н+

 Лактат
Пируват
Лактат
Пируват
Образовавшаяся пировиноградная кислота служит ключевым субстратом в глюконеогенезе.
3.Печень является основным депо гликогена. В гепатоцитах он синтезируется из глюкозо-6-фосфата (Г-6-Ф), УДФ-глюкозы, предшественниками которых могут быть глюкоза, галактоза, фруктоза и другие моносахариды, глицерин, альфа-кетокислоты, аминокислоты (аланин, аспартат, глутамат и другие, называемые гликогенными). Скорость распада гликогена (гликогенолиза) определяется активностью фосфорилазы, которая регулируется различными гормонами и их посредниками – циклическими нуклеотидами.
4.Только в печени и почках обнаруживается глюкозо-6-фосфатаза (схема 1) – фермент, катализирующий следующий гидролиз:
Н2О Фн

 Глюкозо-6-фосфатГлюкоза
Глюкозо-6-фосфатГлюкоза
Глюкозо-6-фосфатаза
Глюкоза крови
Фн Глюкозо-
 6-фосфатаза
6-фосфатаза
 Глюкозо-6-фосфат
Гликоген
Глюкозо-6-фосфат
Гликоген


 УДФ-Галактоза
УДФ-Галактоза
Д инуклеотиды
инуклеотиды
 ПФПУДФ-Глюкоза
ПФПУДФ-Глюкоза

 Полинуклеотиды
Полинуклеотиды

 ГАГ
ГАГ
Гликолиз удф-глюкуроновая гл Мононуклеотиды кислота пг

 АТФ
АТФ
Р
 ибозо-5-фосфат
ПВК
ибозо-5-фосфат
ПВК
 НАДФН+Н+
НАДФН+Н+
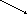

 АТФ ОА
АТФ ОА



 Ацил
КоА Ацетил КоА
Ацил
КоА Ацетил КоА


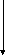 ВЖК
ЦТК
ВЖК
ЦТК

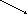
ТАГ АТФ
Фосфолипиды Холестерин Кетоновые тела
Схема 1. Основные пути превращения глюкозы в печени
Фосфорилированная глюкоза, являясь анионом (за счёт фосфата), не способна преодолеть мембрану, однако после гидролиза моносахарид легко диффундирует из клетки в кровь, обеспечивая тем самым стабильность уровня гликемии.
5.Печень – главный орган, регулирующий гомеостаз глюкозы крови за счёт создания динамического равновесия между скоростью синтеза и распада глюкозо-6-фосфата и интенсивностью генеза и расщепления гликогена.
6.Гепатоциты после преобразования Г-6-Ф в УДФ-глюкозу (схема 1) способны использовать последний продукт для получения других моносахаридов и их производных (УДФ-галактозы, УДФ-галактозамина, УДФ-маннозы, УДФ-ацетилгалактозамина, сиаловых кислот и т.д.), которые позднее служат субстратами в синтезе различных гетерополисахаридов, в том числе протеогликанов, липогликанов, гликозаминогликанов.
7.Подобным образом в печени синтезируется УДФ-глюкуроновая кислота, которая, как вышеназванные соединения, может включаться в ГАГи, такие как гиалуроновая кислота, хондроитин-сульфаты А, С, гепарин, гепаран-сульфат. Специфической же функцией УДФ-глюкуроната является участие в инактивации (биотрансформации) различных биологически активных веществ (БАВ): стероидных гормонов, билирубина, лекарственных средств (см. ниже).
8.Только в печени регистрируется активность сорбитол-дегидрогеназы (сорбитол-ДГ), с помощью которой многоатомный спирт – сорбит(ол), окисляется в моносахарид фруктозу. Последняя под действием фруктокиназы или гексокиназы фосфорилируется, что позволяет получившемуся соединению вступить в гликолиз (распад глюкозы). Обнаружение активности сорбитол-ДГ в плазме крови служит своеобразным маркёром патологии печени.
9.В цитоплазме гепатоцитов так же, как и в клетках коры надпочечников, липоцитах, фолликулах щитовидной железы, эритроцитах протекает альтернативный путь окисления глюкозы – пентозофосфатный путь (ПФП). Он не ставит целью синтез АТФ (как в гликолизе), а выполняет следующие функции: а) восстановление НАДФ, который в гепатоцитах необходим в процессе синтеза высших жирных кислот (ВЖК), холестерина (ХС), жёлчных кислот. Кроме того, для гидроксилирования некоторых ядов, лекарственных веществ (амидопирина, морфина, бензофетамина и др.) в микросомах гепатоцитов также используется восстановленный НАДФ; б) образование рибозо-5-фосфата, который в виде фосфорибозилпирофосфата служит одним из субстратов синтеза моно-, ди-, полинуклеотидов.
10.Гипогликемия, развивающаяся после потребления непривычных количеств алкоголя, обусловлена конкурентными взаимоотношениями между необходимостью биотрансформировать этиловый спирт и потребностью глюконеогенеза для поддержания гомеостаза глюкозы крови. Для окисления этанола, так же как и для окисления лактата, требуются значительные величины НАД+; возникает ситуация, характерная для буриданова осла. Чаще она разрешается в пользу алкоголя, провоцируя тем самым гипогликемию (см. схему 2).
11.Глюкозо-6-фосфат (схема 1) через гликолиз и окислительное декарбоксилирование ПВК распадается до ацетил КоА, из которого в печени синтезируются ацилы ВЖК, включающиеся позднее в нейтральные жиры и глицерофосфатиды (судьбу их смотрите ниже).

 Мышцы
Кровь
Печень
Мышцы
Кровь
Печень
 Глюкозо-6-фосфат
Глюкозо-6-фосфат
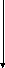
 Гликолиз
Этанол
Гликолиз
Этанол
Л
 актат
Лактат
НАД+
актат
Лактат
НАД+
 Алкоголь-
Алкоголь-
 НАД+
НАДН+Н+
ДГ
НАД+
НАДН+Н+
ДГ


ЛДГ Ацетальдегид

 НАДН+Н+
НАД+
Ацеталь-
НАДН+Н+
НАД+
Ацеталь-
дегид-ДГ
НАДН+Н+
ПВК Ацетат
Схема 2. Механизм конкурентных взаимоотношений