

Рабочий цикл миозиновых мостиков гладкого миоцита
Вхождение актомиозинового комплекса гладкого миоцита в цикл миозиновых головок показан на рис. 709280759 А.

Рис. 709280759. Вхождение актомиозинового комплекса в цикл миозиновых головок (А) и выход из цикла (Б) гладкого миоцита. Киназа ЛЦМ – киназа лёгких цепей миозина.
Инициация сокращения в ответ на увеличение концентрации Ca2+происходит в результате связывания четырёх ионовCa2+ с кальмодулином. Кальций-кальмодулиновый комплекс активирует киназу лёгких цепей миозина, которая фосфорилирует лёгкие цепи на головках миозина. Головках миозина приобретает аффинитет к актну при условии нахождения на ней АДФ и фосфата (продуктов гидролиза АТФ). Начинается цикл миозиновых мостиков.
Расслабление Расслабление скелетного миоцита
Повышенная концентрация Ca2+ в миофибриллярном пространстве сохраняется всего несколько миллисекунд, а затем они «перекачиваются» обратно в цистерны саркоплазматического ретикулума с помощью специального так называемого «кальциевого насоса». Поскольку перекачивание идет против диффузионного градиента, нужна затрата определенной энергии. Ее источником служит расщепление АТФ: на возврат двух ионов Са2+из межфибриллярного пространства в цистерны расходуется одна молекула АТФ.
Ca2+‑АТФаза саркоплазматического ретикулумазакачиваетCa2+из саркоплазмы в цистерны ретикулума, гдеCa2+связывается с кальсеквестрином.
При низкой саркоплазматической концентрации Ca2+тропомиозин закрывает миозинсвязывающие участки и препятствует их взаимодействию с миозином (рис. 709270916 Б).
После смерти, когда содержание АТФ в мышечных волокнах снижается вследствие прекращения её синтеза, головки миозина оказываются устойчиво прикреплёнными к тонкой нити. Это состояние трупного окоченения (rigor mortis)продолжается, пока не наступит аутолиз, после чего мышцы можно растянуть.
У сердечного и гладкого миоцитов Ca2+‑АТФаза саркоплазматического ретикулумазакачиваетCa2+из саркоплазмы в цистерны ретикулума и выкачивает из цитозоля в межклеточное пространство.
У гладкого миоцита миозиновые головки дефосфорилируются фосфатазой легких цепей миозина и головки миозина теряют аффинитет к актину.
Энергетика мышечного сокращения
Единственным прямым (непосредственным) источником энергии для мышечного сокращения служит аденозинтрифосфат (АТФ).
3 химические (энергетические) системы:
фосфагенная, или АТФ-КФ-система;

лактацидная, или гликолитическая;
кислородная, или окислительная.
Рис. 709262218. Три системы синтеза и ресинтеза АТФ при мышечном сокращении: 1 – фосфогенная, 2 – гликолитическая, 3 – окислительная.
19.Режимы и типы сокращений скелетной мышцы. Оптимум и пессимум частоты раздражения скелетной мышцы. Сила и работа мышц. Динамометрия. Закон средний нагрузок.
Раздражение мышечного волокна одиночным пороговым или сверхпороговым стимулом приводит к возникновению одиночного сокращения, которое состоит из нескольких периодов. Первый — латентный период представляет собой сумму временных задержек, обусловленных возбуждением мембраны мышечного волокна, распространением ПД по Т-системе внутрь волокна, образованием инозитолтрифосфата, повышением концентрации внутриклеточного кальция и активации поперечных мостиков. Для портняжной мышцы лягушки латентный период составляет около 2 мс. Второй — период укорочения, или развития напряжения. В случае свободного укорочения мышечного волокна говорят об изотоническом режиме сокращения, при котором напряжение практически не изменяется, а меняется только длина мышечного волокна. Если мышечное волокно закреплено с двух сторон и не может свободно укорачиваться, то говорят об изометрическом режиме сокращения. При данном режиме сокращения длина мышечного волокна не изменяется, в то время как размеры саркомеров меняются за счет скольжения нитей актина и миозина относительно друг друга. В этом случае возникающее напряжение передастся на эластические элементы, расположенные внутри волокна. Эластическими свойствами обладают поперечные мостики миозиновых нитей, актиновые нити, Z-пластинки, продольно расположенная саркоплазматическая сеть и сарколемма мышечного волокна. В опытах на изолированной мышце выявляется растяжение соединительнотканных элементов мышцы и сухожилий, которым передается напряжение, развиваемое поперечными мостиками. В организме человека в изолированном виде изотонического или изометрического сокращения не происходит. Как правило, развитие напряжения сопровождается укорочением длины мышцы — ауксотонический режим сокращения. Третий — период расслабления, когда уменьшается концентрация Са2+ и отсоединяются головки миозина от актиновых филаментов. Полагают, что для одиночного мышечного волокна напряжение, развиваемое любым саркомером, равно напряжению в любом другом саркомере. Поскольку саркомеры соединены последовательно, скорость, с которой происходит сокращение мышечного волокна, пропорциональна числу его саркомеров. Таким образом, при одиночном сокращении скорость укорочения длинного мышечного волокна выше, чем короткого. Величина усилия, развиваемого мышечным волокном, пропорциональна числу миофибрилл в нем. При мышечной тренировке число миофибрилл увеличивается, что является морфологическим субстратом увеличения силы сокращения мышц. Одновременно увеличивается и число митохондрий, повышающих выносливость мышечного волокна при физической нагрузке. В изолированной мышце величина и скорость одиночного сокращения определяются рядом дополнительных факторов. Величина одиночного сокращения в первую очередь будет определяться числом двигательных единиц, участвующих в сокращении. Поскольку мышцы состоят из мышечных волокон с различным уровнем возбудимости, имеется определенная зависимость между величиной стимула и ответной реакцией. Увеличение силы сокращения возможно до определенного предела, после которого амплитуда сокращения остается неизменной при увеличении амплитуды стимула. При этом все мышечные волокна, входящие в состав мышцы, принимают участие в сокращении.
Важность участия всех мышечных волокон в сокращении показана при изучении зависимости скорости укорочения от величины нагрузки. График зависимости скорости сокращения от величины нагрузки приближается к гиперболе. Поскольку сила сокращения эквивалентна нагрузке, становится понятным, что максимальная сила, которая может быть развита мышцей, приходится на очень малые скорости. Штангист может «взять рекордный вес» только при медленных движениях. Напротив, быстрые движения возможны при слабонагруженных мышцах. Изменение силы сокращения наблюдают при ритмической стимуляции скелетных мышц. На рис. 2.15 показаны варианты стимуляции мышцы двумя стимулами. Если второй стимул действует в период рефрактерности мышечного волокна, то он не вызовет повторного мышечного сокращения (рис. 2.15, а)

Если же второй стимул действует на мышцу после окончания периода расслабления, то вновь возникает одиночное мышечное сокращение (рис.2.15, б). При нанесении второго стимула в период укорочения или развития мышечного напряжения происходит суммация двух следующих друг за другом сокращений, и результирующий ответ по амплитуде становится значительно выше, чем при одиночном стимуле. При дальнейшем увеличении частоты стимуляции повторный стимул попадает в период расслабления. В этом случае также возникнет суммация мышечных сокращений, однако будет наблюдаться характерное западение на кривой мышечного сокращения (рис. 2.15, в) — неполная суммация, лежащая в основе зубчатого тетануса. При дальнейшем нарастании частоты стимуляции повторные стимулы будут приходиться на период укорочения, или развития напряжения, вследствие чего происходит полная суммация единичных сокращений, лежащая в основе гладкого тетануса (рис. 2.15, г). Тетанус — сильное и длительное сокращение мышцы. Полагают, что в основе этого явления лежит повышение концентрации кальция внутри клетки, что позволяет осуществляться реакции взаимодействия актина и миозина и генерации мышечной силы поперечными мостиками достаточно длительное время. При тетанусе происходит суммация мышечных сокращений, в то время как ПД мышечных волокон не суммируются.
В естественных условиях одиночные сокращения скелетных мышц не встречаются. Происходит сложение, или суперпозиция, сокращений отдельных нейромоторных единиц. При этом сила сокращения может увели- чиваться как за счет изменения числа двигательных единиц, участвующих в сокращении, так и за счет изменения частоты импульсации мотонейронов. В случае увеличения частоты импульсации будет наблюдаться суммация сокращений отдельных двигательных единиц. Одной из причин увеличения силы сокращения в естественных условиях является частота импульсов, генерируемых мотонейронами. Второй причиной этого служат увеличение числа возбуждающихся мотонейронов и синхронизация частоты их возбуждения. Рост числа мотонейронов соответствует увеличению количества двигательных единиц, участвующих в сокращении, а возрастание степени синхронизации их возбуждения способствует увеличению амплитуды при суперпозиции максимального сокращения, развиваемого каждой двигательной единицей в отдельности. Сила сокращения изолированной скелетной мышцы при прочих равных условиях зависит от исходной длины мышцы. Умеренное растяжение мышцы приводит к тому, что развиваемая ею сила возрастает по сравнению с силой, развиваемой нерастянутой мышцей. Происходит суммирование пассивного напряжения, обусловленного наличием эластических компонентов мышцы, и активного сокращения. Максимальная сила сокращения достигается при размере саркомера 2—2,2 мкм. Увеличение длины саркомера приводит к уменьшению силы сокращения, поскольку уменьшается область взаимного перекрытия актиновых и миозиновых нитей. При длине саркомера 2,9 мкм мышца может развивать силу, равную только 50 % от максимально возможной. В естественных условиях сила сокращения скелетных мышц при их растяжении, например при массаже, увеличивается вследствие работы гамма-эфферентов скелетных мышц.
Работа и мощность мышцы
Поскольку основной задачей скелетных мышц является совершение мышечной работы, в экспериментальной и клинической физиологии оценивают величину работы, которую совершает мышца, и мощность, развиваемую при работе. Согласно законам физики, работа есть энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние: A=FS. Если сокращение мышцы совершается без нагрузки (в изотоническом режиме), то механическая работа равна нулю. Если при максимальной нагрузке не происходит укорочения мышцы (изометрический режим), то работа также равна нулю. В этом случае химическая энергия полностью переходит в тепловую энергию.
Согласно закону средних нагрузок, мьшша может совершать максимальную работу при нагрузках средней величины. При сокращении скелетных мышц в естественных условиях, преимущественно в режиме изометрического сокращения, например при фиксированной позе, говорят о статической работе, при совершении движений — о динамической. Сила сокращения и работа, совершаемая мышцей в единицу времени (мощность), не остаются постоянными при статической и динамической работе. В результате продолжительной деятельности работоспособность скелетных мышц понижается. Это явление называется утомлением. При этом снижается сила сокращений, увеличиваются латентный период сокращения и период расслабления.
Статический режим работы более утомителен, чем динамический. Утомление изолированной скелетной мышцы обусловлено прежде всего тем, что в процессе совершения работы в мышечных волокнах накапливаются продукты окисления — молочная и пировиноградная кислоты, которые снижают возможность генерирования ПД. Кроме того, нарушаются процессы ресинтеза АТФ и креатин фосфата, необходимых для энергообеспечения мышечного сокращения. В естественных условиях мышечное утомление при статической работе в основном определяется неадекватным регионарным кровотоком. Если сила сокращения в изометрическом режиме составляет более 15 % от максимально возможной, то возникает кислородное «голодание» и мышечное утомление прогрессивно нарастает. В реальных условиях необходимо учитывать состояние ЦНС — снижение силы сокращений сопровождается уменьшением частоты импульсации нейронов, обусловленное как их прямым угнетением, так и механизмами центрального торможения. Еще в 1903 г. И.М. Сеченов показал, что восстановление работоспособности утомленных мышц одной руки значительно ускоряется при совершении работы другой рукой в период отдыха первой. В отличие от простого отдыха такой отдых называют активным. Работоспособность скелетных мышц и скорость развития утомления зависят от уровня умственной деятельности: высокий уровень умственного напряжения уменьшает мышечную выносливость.
Динамометрия — метод измерения силы сокращения различных мышечных групп. Для динамометрии существуют различного типа динамометры. Наиболее распространен пружинный динамометр (рис.1). Испытуемый сжимает его кистью вытянутой руки. Сила сжатия указывается стрелкой на специальной шкале. Другая модификация — динамометр Штернберга (рис. 2), имеющий две широкие параллельные ручки, которые испытуемый также сжимает кистью. Существуют ртутные динамометры (рис. 3), в которых сила давления на датчик определяется с помощью ртутного манометра. Разновидность динамометрии — динамография — метод, позволяющий регистрировать силу сокращения мышц графически в виде серии кривых. Этот способ отражает динамику длительного мышечного усилия определенной группы мышц.
20.Физиология гладких мышц: типы, сократительный аппарат, механизм сокращения и расслабление, энергетика.
Гладкие мышцы подразделяют на висцеральные (унитарные) и мультиунитарные (рис. 2.17). Висцеральные гладкие мышцы находятся во всех внутренних органах, протоках пищеварительных желез, кровеносных и лимфатических сосудах, коже. К мультиунитарным относятся ресничная мышца и мышца радужки глаза. Деление гладких мышц на висцеральные и мультиунитарные основано на различной плотности их двигательной иннервации. В висцеральных гладких мышцах двигательные нервные окончания имеются на небольшом количестве гладких мышечных клеток. Несмотря на это, возбуждение с нервных окончаний передается на все гладкие мышечные клетки пучка благодаря плотным контактам между соседними миоцитами — нексусам. Нексусы позволяют потенциалам действия и медленным волнам деполяризации распространяться с одной мышечной клетки на другую, поэтому висцеральные гладкие мышцы сокращаются одномоментно с приходом нервного импульса.
Строение гладких мышц
Гладкие мышцы состоят из клеток веретенообразной формы, средняя длина которых 100 мкм, а диаметр 3 мкм. Клетки располагаются в составе мышечных пучков и тесно прилегают друг к другу. Мембраны прилежащих клеток образуют нексусы, которые обеспечивают электрическую связь между клетками и служат для передачи возбуждения с клетки на клетку. Гладкие мышечные клетки содержат миофиламенты актина и миозина, которые располагаются здесь менее упорядоченно, чем в волокнах скелетной мышцы. Саркоплазматическая сеть в гладкой мышце менее развита, чем в скелетной.
Иннервация гладких мышц
Висцеральная гладкая мышца имеет двойную — симпатическую и парасимпатическую иннервацию, функция которой заключается в изменении деятельности гладкой мышцы. Раздражение одного из вегетативных нервов обычно увеличивает активность гладкой мышцы, стимуляция другого — уменьшает. В некоторых органах, например кишечнике, стимуляция адренергических нервов уменьшает, а холинергических увеличивает мышечную активность; в других, например сосудах, норадреналин усиливает, а АХ снижает мышечный тонус. Строение нервных окончаний в гладкой мышце отличается от строения нервно-мышечного синапса скелетной Мышцы. В гладкой мышце нет концевых пластинок и отдельных нервных окончаний. По всей длине разветвлений адренергических и холинергических нейронов имеются утолщения, называемые варикозами. Они содержат гранулы с медиатором, который выделяется из каждой варикозы нервных волокон. Таким образом, по ходу следования нервного волокна могут возбуждаться или тормозиться многие гладкие мышечные клетки. Клетки, лишенные непосредственных контактов с варикозами, активируются потенциалами действия, распространяющимися через нексусы на соседние клетки. Скорость проведения возбуждения в гладкой мышце невелика и составляет несколько сантиметров в секунду. Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в виде отдельных волн деполяризации. При повторной стимуляции эти потенциалы суммируются и по достижении пороговой величины возникает ПД. Тормозящее влияние адренергических или холинергических нервов проявляется в виде отдельных волн гиперполяризации, называемых тормозными постсинаптическими потенциалами (ТПСП). При ритмической стимуляции ТПСП суммируются. Возбуждающие и тормозные постсинаптические потенциалы наблюдаются не только в мышечных клетках, контактирующих с варикозами, но и на некотором расстоянии от них. Это объясняется тем, что постсинаптичсские потенциалы передаются от клетки к клетке через нексусы или посредством диффузии медиатора из мест его выделения.
Автоматия. ПД гладких мышечных клеток имеют авторитмический (пейсмекерный) характер, подобно потенциалам проводящей системы сердца. Пейсмекерные потенциалы регистрируются в различных участках гладкой мышцы. Это свидетельствует о том, что любые клетки висцеральных гладких мышц способны к самопроизвольной автоматической активности. Автоматия гладких мышц, т.е. способность к автоматической (спонтанной) деятельности, присуща многим внутренним органам и сосудам.
Реакция на растяжение. Уникальной особенностью висцеральной гладкой мышцы является се реакция на растяжение. В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение уменьшает МП клеток, увеличивает частоту ПД и в конечном итоге — тонус гладких мышц. В организме человека это свойство гладких мышц служит одним из способов регуляции двигательной деятельности внутренних органов. Например, при наполнении желудка происходит растяжение его стенки. Увеличение тонуса стенки желудка в ответ на его растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления, является основным фактором миогенной саморегуляции тонуса сосудов. Наконец, растяжение мускулатуры матки растущим плодом служит одной из причин начала родовой деятельности.
Связь возбуждения с сокращением. Изучать соотношения между электрическими и механическими проявлениями в висцеральной гладкой мышце труднее, чем в скелетной или сердечной, так как висцеральная гладкая мышца находится в состоянии непрерывной активности. В условиях относительного покоя можно зарегистрировать одиночный ПД. В основе сокращения как скелетной, так и гладкой мышцы лежит скольжение актина по отношению к миозину, где Са2+ выполняет триггерную функцию (рис. 2.18). В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы: прежде чем миозин гладкой мышцы сможет проявлять свою АТФазную активность, он должен быть фосфорилирован. Фосфорилирование и дефосфорилирование миозина наблюдается и в скелетной мышце, но в ней процесс фосфорилирования не является обязательным для активации АТФазной активности миозина. Механизм фосфорилирования миозина гладкой мышцы осуществляется следующим образом: Са2+ соединяется с кальмодулином (кальмодулин — рецептивный белок для Са2+). Возникающий комплекс активирует фермент — киназу легкой цепи миозина, который в свою очередь катализирует процесс фосфорилирования миозина. Затем происходит скольжение актина по отношению к миозину, составляющее основу сокращения. Отмстим, что пусковым моментом для сокращения гладкой мышцы является присоединение Са2+ к кальмодулину, в то время как в скелетной и сердечной мышцах пусковым моментом является присоединение Са2+ к тропонину.
21.Физиологические свойства миокарда. Автоматия сердца. Проводящая система сердца, её функциональные особенности.
Все мышечные клетки сердца (кардиомиоциты или миокардиоциты) можно разделить на 3 типа:
рабочие
атипичные
секреторные.
У каждого типа кардиомиоцита своя функция
рабочих – сократимость, обеспечивающая нагнетательную функцию сердца
атипичные – проводимость, обеспечивающая оптимальный характер распространения возбуждения по миокарду
секреторные – инкреторная.
Физиологические свойства
Как всем клеткам возбудимых тканей миокардиоцитам присущи следующие четыре свойства: возбудимость, проводимость, автоматизм и способность формировать специфический ответ.
Для рабочих миокардиоцитов специфическим ответом является сокращение (сократимость).
Для секреторных миокардиоцитов специфическим ответом является секреция гормона.
Изменение физиологических свойств миокарда (типы влияний на свойства миокарда
Влияние веществ и других факторов
на возбудимость миокарда носит название батмотропия,
на автоматизм — хронотропия,
на проводимость — дромотропия,
на сократимость — инотропия.
Как проявляются эффекты?
Батмотропный – изменением порога раздражения:
положительный – уменьшением,
отрицательный – увеличением.
Дромотропный – изменением скорости проведения возбуждения:
положительный – увеличением,
отрицательный – уменьшением.
Хронотропный – изменением частоты сердечных сокращений:
положительный – увеличением,
отрицательный – уменьшением.
Инотропный – изменением силы сокращения:
положительный – увеличением,
отрицательный – уменьшением.
Типы инотропных влияний на миокард
Действие инотропных факторов может быть направлено на регуляцию двух процессов, определяющих силу сокращения:
на увеличение числа взаимодействующих мостиков (например, при увеличении концентрации Са2+).
на интенсификацию процессов, протекающих в каждом отдельном пункте взаимодействия, т.е. на кинетические свойства отдельных мостиков (например, при действии катехоламинов, сердечных гликозидов, симпатической стимуляции).
Проводящая
система сердца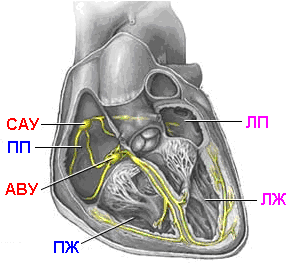
САУ – синоатриальный узел,
ПП – правое предсердие,
ЛП – левое предсердие,
АВУ – атриовентрикулярный узел,
ПЖ – правый желудочек,
ЛЖ – левый желудочек.
По функциональным особенностям, по специфике морфологии и ультраструктуры проводящую систему высших позвоночных и человека делят на следующие отделы :
Узловую ткань
Синусовый узел (Киса-Фляка)
атриовентрикулярный узел (Ашоффа-Тавара);
Собственно проводящие пути
Проводящие пути предсердий
Межпредсердный путь Бахмана
Межузловые проводящие пути предсердий
Бахмана
Венкебаха
Тореля
Пучок Гиса,
Ножки пучка Гиса
Правая
Левая
Передняя ветвь
Задняя ветвь
Субэндокардиальная сеть волокон Пуркинье.
Синусовый (синоатриальный) узелрасположен в верхней части правого предсердия в устье верхней полой вены. Длина узла составляет 1,5 — 2,5 см, ширина — 0,4 — 0,7 см. Различают головку синусового узла, расположенную субэпикардиально, и хвост, располагающийся субэндокарднально.
В синусовом узеле выделяют четыре морфологических типа клеток: пейсмекеры(Р-клетки), недифференцированные, промежуточные, волокна Пуркинье.
Пейсмекеры расположенные главным образом в центре узла. Недифференцированные клетки миокарда находятся преимущественно в периферических отделах узла. Волокна Пуркинье располагаются по краям синусового узла и подходят к предсердным проводящим путям. Промежуточные клетки обладающие морфологическими признаками пейсмекеров и недифференцированных клеток.
Кровоснабжение синусового узла осуществляет ramus cristae terminalis, артерии, отходящей от правой (иногда от левой) венечной артерии.
Проводящие
пути предсердий (вид сверху и сзади)
1 – межпредсердный путь Бахмана,
2 – передний межузловой путь Бахмана,
3 – средний межузловой путь Венкебаха,
4 – задний межузловой путь Тореля.
СУ – синоатриальный узел,
АВУ – атриовентрикулярный узел.
ВПВ – верхняя полая вена.
Атриовентрикулярный узел
Выделяют четыре типа клеток атриовентрикулярного узла, аналогичных клеткам синусового узла: пейсмекеры, переходные и сократительные клетки, клетки Пуркинье.
Пейсмекеров немного и он располагаются главным образом в области перехода атриовентрикулярного узла в пучок Гиса. Переходные клетки составляют основную массу атриовентрикулярного узла. Клетки сократительного миокарда, располагающиеся главным образом у атрионодального края.