

11.Синапс: строение, классификация, синаптические токи, процесс передачи импульса.
Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном. Он, исследуя деятельность ЦНС, предположил еще в 1897 г., что нейроны между собой сообщаются с помощью специального механизма, который он и назвал синаптическим. Последующие годы подтвердили эту идею.
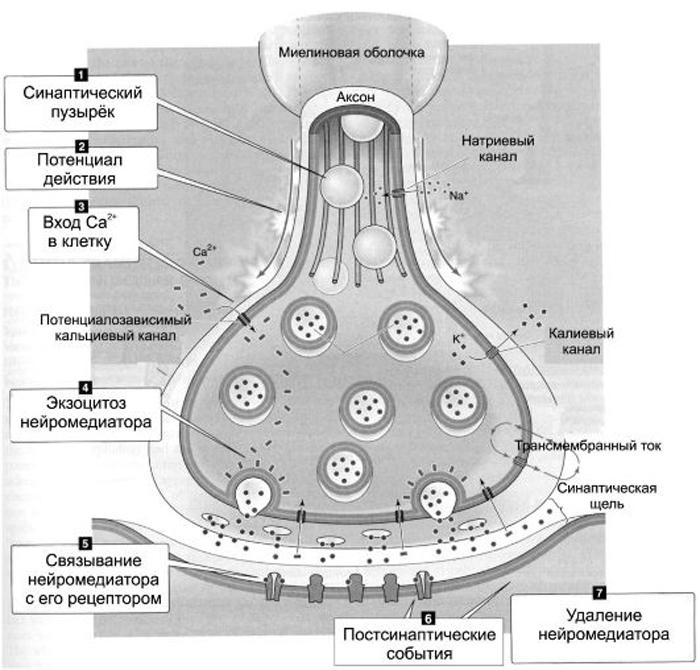
Синаптическая передача
Классификация синапсов
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Все синапсы ЦНС можно классифицировать следующим образом.
1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
3. По конечному эффекту: тормозные и возбуждающие.
4. По механизму передачи сигнала: электрические, химические, смешанные.
5. Химические синапсы можно классифицировать:
а) по форме контакта – терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона);
б) по природе медиатора – холинергические (медиатор – ацетилхолин, АХ), адренергическис (медиатор – норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор – гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор – пептиды, например, вещество Р), пуринергические (медиатор – АТФ).
Электрические синапсы. В настоящее время признают, что в ЦНС имеются электрические синапсы. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом – медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, постсинаптический элемент, который ограничен постсипаптической мембраной, а также внесинаптическую область и синаптическующель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка – это синапс между нейронами, концевая пластинка – это постсинаптическая мембрана мионеврального синапса, моторная бляшка – это пресинаптичсское окончание аксона на мышечном волокне.
Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию мембраны, которая включает кальциевый насос, и ионы кальция поступают в синапс. Попадая в цитоплазму синаптического окончания, кальций связывается с белками оболочки синаптических везикул (пузырьков, в которых хранятся медиаторы), что приводит к выделению медиаторов в синаптическую щель, которая отделяет мембрану одного нейрона от мембраны другого. Так возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический, т. е. каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества - медиатора. Далее молекулы медиатора связываются со специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул (которые подходят как "ключ к замку"). Рецепторы - белковые структуры, которые являются интегральными белками плазматической мембраны. Они синтезируются в рибосомах эндоплазматическогоретикулюма клетки, затем встраиваются в мембраны. Функциональная активность синапса зависит от количества рецепторов, а также от их сродства к медиатору (лиганду). Искусственно созданные лиганды, обладающие высоким специфическим (т. е. действующим только на определенный тип либо подтип рецептора) сродством к рецепторам, способны вызывать такой же функциональный ответ клетки, как и нативные медиаторы. Рецептор состоит из двух частей. Одну можно назвать "узнающим центром", другую - "ионным каналом". Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) постсинаптсинаптический потенциала (ВПСП) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал (ТПСП). На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона (на последних находится до нескольких тысяч тормозных и возбудительных синапсов).
После каждого проведения импульса медиаторы разрушаются специфическими ферментами либо происходит их обратный захват в пресинаптическое окончание. В синапсах существует широко распространенное в физиологии явление обратной связи: медиатор из синаптической щели может взаимодействовать с рецепторами на пресинаптической мембране (ауторецепторами), что приводит к прекращению выброса медиатора. Активность синапса может модулироваться действием модулирующих нейромедиаторов, рецепторы которых располагаются на аксоне или даже дендритах и теле синапса. Если через синапс проходит много импульсов, то рецепторы могут уменьшить свою чувствительность к нейромедиатору (это свойство рецепторов обеспечивает процессы адаптации на нейрональном уровне). Важно помнить, что между химической природой синапса и знаком его синаптического действия (возбуждающий или тормозной) нет однозначной зависимости: один и тот же медиатор может оказывать как тормозное, так и возбуждающее действие. Знак синаптического действия определяется свойствами постсинаптической мембраны, т. е. составом рецепторов, которые могут разным образом реагировать с медиатором и контролировать проводимость разных ионных каналов.