

1) Каждый наследственный признак определяется отдельным наследственным фактором, задатком; в современном представлении эти задатки соответствуют генам;
2) Гены сохраняются в чистом виде в ряду поколений, не утрачивая своей индивидуальности: это явилось доказательством основного положения генетики: ген относительно постоянен;
3) Оба пола в равной мере участвуют в передаче своих наследственных свойств потомству;
4) Редупликация равного числа генов и их редукция в мужских и женских половых клетках; это положение явилось генетическим предвидением существования мейоза;
5) Наследственные задатки являются парными, один – материнский, другой – отцовский; один из них может быть доминантным, другой – рецессивным; это положение соответствует открытию принципа аллелизма: ген представлен минимум двумя аллелями.
К законам наследованияотносятся закон расщепления наследственных признаков в потомстве гибрида и закон независимого комбинирования наследственных признаков. Эти два закона отражают процесс передачи наследственной информации в клеточных поколениях при половом размножении. Их открытие явилось первым фактическим доказательством существования наследственности как явления.
Законы наследственностиимеют другое содержание, и они формулируются в следующем виде:
Первый закон – закон дискретной (генной) наследственной детерминации признаков; он лежит в основе теории гена.
Второй закон – закон относительного постоянства наследственной единицы – гена.
Третий закон – закон аллельного состояния гена (доминантность и рецессивность).
То, что законы Менделя связаны с поведением хромосом при мейозе, было обнаружено в начале ХХ века во время повторного открытия законов Менделя сразу тремя группами ученых независимо друг от друга. Как вам уже известно, особенность мейоза заключается в том, что число хромосом в клетке уменьшается вдвое, хромосомы могут меняться своими частями при мейозе. Такая особенность характеризует ситуацию с жизненным циклом у всех эукариот.
Для того, чтобы проверить предположение о наследовании задатков в таком виде, как мы уже говорили, Мендель провел также скрещивание потомков первого поколения, имеющие желтые семена с родительскими зелеными (рецессивными). Скрещивание на рецессивный организм он назвал анализирующим. В результате он получил расщепление один к одному: (Аа х аа = Аа + Аа + аа + аа). Таким образом, Мендель подтвердил предположение, что в организме первого поколения есть задатки признаков каждого из родителей в соотношении 1 к 1. Состояние, когда оба задатка признака одинаковы, Мендель назвал гомозиготным, а когда разные – гетерозиготным.

Мендель учитывал результаты, полученные на тысячах семян, то есть он проводил статистические исследования, которые отражают биологическую закономерность. Открытые им самые законы будут действовать и на других эукариотах, например грибах. Здесь показаны грибы, у которых четыре споры, получаемые в результате одного мейоза, остаются в общей оболочке. Анализирующее скрещивание у таких грибов приводит к тому, в одной оболочке присутствуют 2 споры с признаком одного родителя и две с признаком другого. Таким образом, расщепление 1:1 в анализирующем скрещивании отражает биологическую закономерность расщепления задатков одного признака в каждом мейозе, которая будет выглядеть как закономерность статистическая, если все споры смешать.
То, что у родителей были разные состояния одного признака, говорит о том, что задатки к развитию признака могут как-то меняться. Эти изменения называются мутациями. Мутации бывают нейтральными: форма волос, цвет глаз и др. Некоторые мутации приводят к изменениям, нарушающим нормальное функционирование организма. Это коротконогость у животных (крупный рогатый скот, овцы и др.), безглазость и бескрылость у насекомых, бесшерстность у млекопитающих, гигантизм и карликовость.
Некоторые мутации могут быть и безвредными, например бесшерстность у людей, хотя все приматы имеют волосяной покров. Но иногда встречаются изменения интенсивности волосяного покрова на теле и у людей. Н.И.Вавилов назвал такое явление законом гомологических рядов наследственной изменчивости: то есть признак, типичный только для одного из двух родственных видов, может быть обнаружен с какой-то частотой и у особей родственного вида.
На фотографии показано , что мутации могут быть достаточно заметными, мы видим негритянскую семью, в которой родился белый негр – альбинос. У него дети, скорее всего, будут пигментированными, поскольку мутация эта рецессивная, а частота ее встречаемости низка.

Мы говорили до этого о признаках, которые проявляются полностью. Но это не для всех признаков так. Например, фенотип гетерозигот может быть промежуточным между доминантным и рецессивным признаком родителей. Так, окраска плода у баклажан в первом поколении меняется с темно-синей на менее интенсивную фиолетовую. При этом во втором поколении расщепление по наличию окраски осталось 3:1, но если учитывать интенсивность окраски расщепление стало 1:2:1 (цвет темно-синий - АА, фиолетовый - 2Ааи белый -аа, соответственно) В данном случае видно, что проявление признака зависит от дозы доминантного аллеля. Расщепление по фенотипу соответствует расщеплению по генотипу: классыАА,Ааиаа, в соотношении 1:2:1.

Еще раз выделим роль Менделя в развитии науки. Никто до него не размышлял, что вообще могут существовать задатки признаков. Считалось, что в каждом из нас сидит маленький человечек, внутри его – еще маленький человечек и т.д. Зачатие имеет к его появлению какое-то отношение, но по механизму, готовый маленький человечек уже присутствует с самого начала своего роста. Такими были доминирующие представления, у которых, безусловно, был недостаток – по этой теории, при большом числе поколений гомункулус должен был получиться по размеру меньше элементарной частицы, но тогда про частицы еще не знали J.
Откуда Мендель знал, какой признак является доминантным, а какойрецессивным? Ничего такого он не знал, просто взял некоторый принцип организации опыта. Удобно, что признаки, за которыми он наблюдал, были разными: рост, размер, цвет цветка, цвет боба и т.д. У него не было априорной модели механизма наследования, он вывел ее из наблюдения за передачей признака в поколениях. Еще одна особенность его метода. Он получил, что доля особей с рецессивным признаком во втором поколении составляет четверть от всего потомства. То есть вероятность того, что данная горошина зеленая – 1/4. Допустим получилось в среднем по 4 горошины в одном стручке. Будет ли в каждом стручке (это потомство от двух и только от двух родителей) 1 горошина зеленая и 3 желтых? Нет. Например, вероятность того, что там будет 2 зеленых горошины равна 1/4 х 1/4 = 1/16, а того, что все четыре зеленые – 1/256. То есть, если взять кучу бобов, с четырьмя горошинами в каждом, то у каждой 256-ой все горошины будут с рецессивными признаками, то есть зелеными. Мендель анализировал потомство множества одинаковых пар родителей. О скрещивании было рассказано, потому что они показывают, что законы Менделя проявляются как статистические, а в основе имеют биологическую закономерность – 1:1. То есть гаметы разных типов в КАЖДОМ мейозе у гетерозиготы образуются в равном соотношении – 1:1, а закономерности проявляется статистически, поскольку анализируются потомки сотен мейозов – Мендель анализировал более 1000 потомков в скрещивании каждого типа.
Сначала Мендель исследовал наследование одной пары признаков. Затем он задался вопросом, что будет происходить, если одновременно наблюдать за двумя парами признаков. Выше на рисунке, в правой части проиллюстрировано такое исследование по дум парам признаков - цвету горошин и форме горошин.
Родители одного типа давали при самоопылении горошины желтые и круглой формы. Родители другого типа давали при самоопылении горошины зеленые и морщинистой формы. В первом поколении он получил все горошины желтые, а по форме - круглые. Получившееся расщепление во втором поколении удобно рассмотреть с помощью решетки Пенета. Получили расщепление по признакам 9:3:3:1 (желтые и круглые: желтые и морщинистые: зеленые и круглые: зеленые и морщинистые). Расщепление по каждой паре признаков происходит независимо друг от друга. Соотношение 9жк + 3жм + 3зк + 1зм соответствует независимой комбинации результатов двух скрещиваний (3ж + 1з) х (3к + 1 м). То есть и задатки признаков этих пар (цвет и форма) комбинируются независимо.
Посчитаем, сколько разных фенотипических классов мы получили. У нас было 2 фенотипических класса: желтые и зеленые; и по другому признаку 2 фенотипических класса: круглые и морщинистые. А всего будет 2*2=4 фенотипических класса, что мы и получили выше. Если рассматривать три признака, то фенотипических классов будет 23=8 классов. Мендель доходил до дигибридных скрещиваний. Задатки всех признаков, к счастью Менделя, находились у гороха на разных хромосомах, а всего хромосом у гороха – 7 пар. Поэтому, оказалось, что он взял признаки, которые комбинировались независимо в потомстве.
У человека 23 пары хромосом. Если рассмотреть какой-то один гетерозиготный признак для каждой хромосомы, может у человека может наблюдаться 223 ~ 8*106 фенотипических классов в потомстве одной супружеской пары. Как упоминалось на первой лекции, каждый из нас содержит между папиными и мамиными хромосомами порядка 1 различия на 1000 позиций, то есть всего порядка миллиона различий между папиными и мамиными хромосомами. То есть каждый из нас является потомком миллионногибридного скрещивания, при котором число фенотипических классов составляет 21000000. Практически это число фенотипических классов в потомстве одной пары не реализуется, потому что хромосом у нас всего 23, а не миллион. Получается, что 8*106 – это нижний предел величины возможного разнообразия в потомстве данной супружеской пары.Исходя из этого, можно понять, что не может быть двух абсолютно одинаковых людей.Вероятность мутации данного нуклеотида в ДНК за одно поколение составляет около 10-7- 10-8, то есть на весь геном (3*109) получится около 100 изменений de novo между родителем и ребенком. А всего отличий в папиной половинке вашего генома от маминой половинки – около 1 000 000. Это значит, что старые мутации в вашем геноме гораздо более частые, чем вновь возникшие (в 10 000 раз).
Также Мендель проводил анализирующее скрещивание - скрещивание с рецессивной гомозиготой. У потомка первого поколения комбинация генов имеет вид АаВb. Если скрестить его с представителем с полностью рецессивным набором генов (aabb), то получится четыре возможных класса, которые будут находиться всоотношении 1:1:1:1, в отличие от рассмотренного выше скрещивания, когда мы получили расщепление 9:3:3:1.

Ниже показаны некоторые статистические критерии – какие соотношения чисел следует считать соответствующими ожидаемым, скажем, 3:1. Например, для 3:1 - из четырехсот горошин вряд ли получится точно 300 к 100. Если получится, к примеру, 301 к 99, то это отношение наверное можно считать равным 3 к 1. А 350 к 50 уже, наверное, не равно 3 к 1. Статистический тест хи-квадрат (χ2) используетсядля проверки гипотезы соответствия наблюдаемого распределения ожидаемому. Произносится эта греческая буква в русском языке как «хи», а в английском - как «чи» (chi).
Величина χ2 рассчитывается как сумма квадратов отклонений наблюдаемых величин от ожидаемой, деленных на ожидаемую величину. Затем по специальной таблице для данного значения χ2 находят величину вероятности того, что такое различие между наблюдаемой и ожидаемой величиной является случайным. Если вероятность оказывается меньше 5% то отклонение считается не случайным (цифра в пять процентов выбрана по договоренности).

Всегда ли будет проявляться какой-либо наследственно предопределенный признак? Ведь это предположение по умолчанию лежит в основе интерпретации данных полученных Менделем. Оказывается, это может зависеть от многих причин. Есть такая наследуемая черта у человека – шестипалость. Хотя у нас, как и у всех позвоночных, пальцев в норме пять. Вероятность проявления задатка признака в виде наблюдаемого признака (здесь – шестипалость) может быть меньше 100%. На фотографии у человека на обеих ногах по 6 пальцев. А у его близнеца этот признак не обязательно проявится. Доля индивидов с данным генотипом, у которых проявляется соответствующий фенотип, была названа пенетрантностью (этот термин ввел российский генетик Тимофеев-Ресовский).
В некоторых случаях шестой палец может быть просто обозначен некоторым кожным приростом. Степень выраженности признака у индивида Тимофеев-Ресовский предложил называть экспрессивностью.
Особенно ясно не 100% связь генотипа с фенотипомпрослеживается при исследовании идентичных близнецов. Генетическая конституция у них один в один, а признаки у них совпадают в разной степени. Ниже представлена табличка, в которой представлено совпадение признаков для близнецов идентичных и неидентичных. В качестве признаков в этой таблице взяты различные болезни.

Признак, который присутствует у большинства особей в естественных условиях обитания, называется диким типом.Наиболее распространенный признак часто оказывается доминантным. Такая связь может иметь приспособительное значение, полезное для вида. У человека доминантными признаками являются, к примеру, черные волосы, темные глаза, кудрявые волосы. Кстати, поскольку соответствующие гены находятся на разных хромосомах, то можетполучиться кудрявый негр, который будет блондином - ничто это не запрещает.
Почему так получается, что в при моногибридном скрещивании трем генотипическим классам в потомстве второго поколения соответствует в некоторых случаях три фенотипических класса (баклажаны синие фиолетовые и белые), а в другом случае – два класса (желтая или зеленая горошина)? Почему в одном случае проявление доминантного признака неполное, а в другом – полное? Можно провести аналогию с фотопленкой. В зависимости от количества света, кадр может получиться совсем прозрачным, серым и совсем черным. То же самое – с генами. Например, есть у кукурузы ген Y, который определяет образование витамина А. Когда доза гена Y на клетку растет от одного до трех, то линейно изменяется активность фермента, который он кодирует и, в данном случае, усиливается образование витамина А и окраска зерна. (У кукурузы основная часть зерна – эндосперм. В каждой клетке эндосперма три генома – два от мамы и один от папы). То есть, многие признаки зависят от дозы аллеля количественно. Чем больше копий аллеля нужного типа, тем больше будет величина контролируемого им признака. Такая связь постоянно используется в биотехнологии.

Мендель мог благополучно свои законы и не открыть. Исследования на горохе позволилиМенделю открыть свои законы, потому что горох - самоопыляемое растение, а потому без принуждения – гомозиготный. При самоопылении доля гетерозигот уменьшается пропорционально двум в степени номера поколения. В этом заключалось везение Менделя– если бы доля гетерозигот была большой, то никаких бы закономерностейне наблюдалось. Когда он затем взял перекрестные опылители, то закономерности нарушились, что сильно расстроило Менделя, потому что он подумал, что открыл нечто частное. Оказалось, что нет.

Выше было рассказано о наследовании признаков качественных, а обычно большинство признаков - количественные.Их генетический контроль достаточно сложен. Количественные признаки описываются через среднюю величину значения признака и размахом варьирования, которая называется нормой реакции. И величина средней, и норма реакции - это видоспецифические показатели, которые зависят как от генотипа, так и от условий среды. К примеру, продолжительность жизни человека. Хоть в Библии инаписано, что пророки жили по 800 лет, но сейчас ясно, что больше 120-150 лет никто не живет. А, мышь, например, живет два года, хотя она тоже млекопитающее. Наш рост, наш вес – это все количественные признаки. Нет людей 3-4 метрового роста, хотя слоны, к примеру, есть. У каждого вида своя средняя по каждому количественному признаку и свой размах его варьирования.
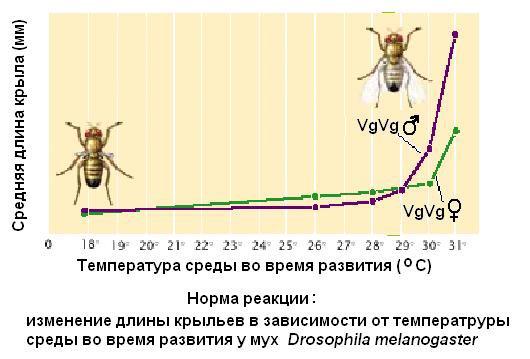
Закономерности наследования открыты при исследовании качественных признаков.
Большинство наших признаков – количественные.
Величины значений признаков в представительной выборке особей данного вида характеризуются определенной средней и широтой ее варьирования, которая называется нормой реакции и зависит как от генотипа, так и от условий формирования признака.
Деление клетки (митоз, мейоз и их отличиях) и индивидуальное развитие (образовании гамет и оплодотворения и краткий обзор эмбрионального развития).
Клетка в своей жизни проходит разные состояния: фазу роста и фазы подготовки к делению и деления. Клеточный цикл – переход от деления к синтезу веществ, составляющих клетку, а затем опять к делению – можно представить на схеме в виде цикла, в котором выделяют несколько фаз.

После деления клетка вступает в фазу синтеза белков и роста, эту фазу называют G1. Часть клеток из этой фазы переходит в фазу G0, эти клетки функционируют и потом погибают без деления (например, эритроциты). Но большинство клеток, накопив необходимые вещества и восстановив свой размер, а иногда и без изменения размеров после предыдущего деления, начинают подготовку к следующему делению. Эта фаза называется фаза S – фаза синтеза ДНК, затем, когда хромосомы удвоились, клетка переходит в фазу G2 – фазу подготовки в митозу. Затем происходит митоз (деление клетки), и цикл повторяется заново. Фазы G1, G2, S вместе называются интерфазой (т.е. фазой между делениями клетки).

Жизнь клетки и переход от одной фазы клеточного цикла к другойрегулируется изменениемконцентраций белковциклинов , как это показано на рисунке.

При подготовке к делению происходит репликация ДНК, на каждой хромосоме синтезируется ее копия. Пока эти хромосомы после удвоения не расходятся, каждая хромосома в этой паре называется хроматидой. После репликации ДНК конденсируется, хромосомы приобретают более компактную укладку, и в таком состоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в большей степени расплетены. Понятно, что в конденсированном состоянии им трудно функционировать. Хромосома имеет вид в виде буквы Х только во время одной из стадий митоза. Раньше считалось, что между делениями клетки хромосомная ДНК ( хроматин ) находится в полностью расплетенном состоянии, но сейчас выясняется, что структура хромосом достаточно сложная и степень деконденсации хроматина между делениями не очень велика.

Процесс деления, при котором исходно диплоидная клетка дает две дочерние, также диплоидные, клетки, называетсямитозом. Имеющиеся в клетке хромосомы удваиваются, выстраиваются в клетке, образуя митотическую пластинку, к ним прикреплены нити веретена деления, которые растягиваются к полюсам клетки и клетка делится, образуя две копии исходного набора.
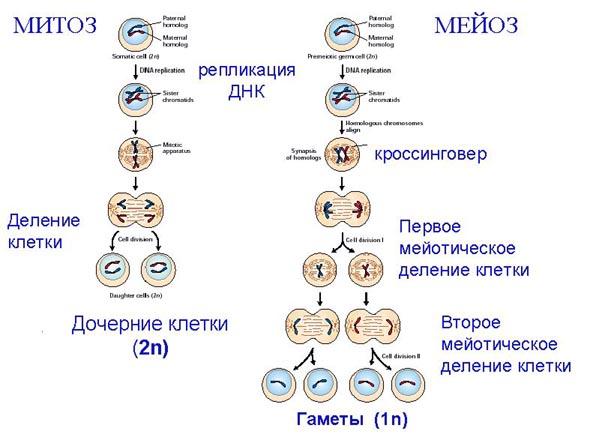
При образовании гамет, т.е. половых клеток – сперматозоидов и яйцеклеток – происходит деление клетки, называемое мейозом. Исходная клетка имеет диплоидный набор хромосом, которые затем удваиваются. Но, если при митозе в каждой хромосоме хроматиды просто расходятся, то при мейозе хромосома (состоящая из двух хроматид) тесно переплетается своими частями с другой, гомологичной ей хромосомой (также состоящей из двух хроматид), и происходит кроссинговер - обмен гомологичными участками хромосом. Затем уже новые хромосомы с перемешанными «мамиными» и «папиными» генами расходятся и образуются клетки с диплоидным набором хромосом, но состав этих хромосом уже отличается от исходного, в них произошла рекомбинация . Завершается первое деление мейоза, и второе деление мейоза происходит без синтеза ДНК, поэтому при этом делении количество ДНК уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидных клетки. Фазы деления клетки, которые следуют за интерфазой, называются профаза, метафаза, анафаза, телофаза и после деления опять интерфаза.

При мейозе фазы называется также, но указывается к какому делению мейоза она относится. Кроссинговер – обмен частями между гомологичными хромосомами – происходит в профазе первого деления мейоза (профаза I), которая включает следующие этапы: лептотена, зиготена, пахитена, диплотена, диакинез. Процессы, происходящие при этом в клетке..


Краткий обзор этапов гаметогенеза .Гаметогенез подразделяется насперматогенез (процесс образования сперматозоидов у самцов) иоогенез (процесс образования яйцеклетки). По тому, что происходит с ДНК, эти процессы практически не отличаются: одна исходная диплоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с цитоплазмой, эти процессы кардинально различаются.
В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем для развития зародыша, поэтому яйцеклетка – это очень крупная клетка, и когда она делится, цель – сохранить питательные вещества для будущего зародыша, поэтому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы цитоплазмы и при этом избавиться от ненужного генетического материала, от цитоплазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы, но позволяют поделить хромосомный набор. Полярные тельца отделяются при первом и втором делении мейоза (

Исходная клетка, из которой в последствии образуется зрелая яйцеклетка, называется ооцитом первого порядка. После деления из него образуется ооцит второго порядка и первое полярное тельце. Затем происходит второе деление мейоза, в результате образуетсягаплоидный оотид и второе полярное тельце. Первое полярное тельце за это время тоже успевает поделиться, таким образом всего получается три гаплоидных полярных тельца. Воотидепроисходят некоторые процессы созревания и он превращается в яйцеклетку. Она содержащая почти всю цитоплазму исходного ооцита, но гаплоидный набор хромосом. Эти хромосомы уже прошли рекомбинацию, т.е. если исходно клетки содержат одну хромосому от мамы, одну от папы, то в зрелой яйцеклетке в каждой хромосоме чередуются куски, полученные от одного и второго родителя.
При сперматогенезе цитоплазма исходного сперматоцита первого порядкаделится (первое деление мейоза) поровну между клетками, давая сперматоциты второго порядка.Второе деление мейозаприводит к образованию гаплоидных сперматоцитов второго порядка. Затем происходит созревание без деления клетки, большая часть цитоплазмы отбрасывается, и получаются сперматозоиды, содержащие гаплоидный набор хромосом очень мало цитоплазмы. Ниже представлена фотография сперматозоида человека и схема его строения.
Сперматозоиды животных имеют одинаковое принципиальное строение, но могут отличаться формой и размером. Сперматозоид имеет головку, в которую плотно упакована ДНК. Головка сперматозоида окружена очень тонким слоем цитоплазмы. На ее переднем конце находится структура, называемая акросомой. Эта структура содержит ферменты, позволяющие сперматозоиду проникнуть через оболочку яйцеклетки. Сперматозоид имеет хвостик. Часть хвостика, прилегающая к головке ("шейка"), окружена митохондриями. Они необходимы, чтобы обеспечить биение хвостика и движение сперматозоида в желательном ему направлении. На сперматозоиде имеется для выбора направления движения хеморецепторы, сходные с обонятельными клетками.

Созревание спермиев происходит в семенных канальцах тестикул. При превращении исходной клетки, сперматогония, в сперматоцит, сперматиды и зрелый сперматозоид происходит перемещение клетки от базальной мембраны семенного канатика к его полости. После созревания сперматозоиды отделяются, попадая в просвет семенных канальцев, и готовы к движению в поисках яйцеклетки и оплодотворению. Процесс созревания длится примерно три месяца.У млекопитающих у особей мужского пола процесс созревания сперматозоидов – сперматогенез – начинается с возраста половой зрелости и продолжается затем до глубокой старости.

Существенно отличается процесс созревания яйцеклетки – оогенез. Во время эмбрионального развития млекопитающих возникает большое количество яйцеклеток, и к рождению самки в ее яичниках уже находится порядка 200-300 тысяч яйцеклеток, остановившихся на первой стадии деления мейоза. В период полового созревания яйцеклетки начинают реагировать на половые гормоны, Регулярные циклические изменения гормонов впоследствии вызывают созревание яйцеклетки, обычно одной, иногда двух или больше. Когда для лечения бесплодия женщине делают инъекции половых гормонов, чтобы индуцировать созревание яйцеклеток, избыток этих гормонов может привести к созреванию нескольких яйцеклеток, и как следствие этого – многоплодной беременности. Яйцеклетка созревает в пузырьке, называемом фолликулом.

За всю жизнь у женщин современных индустриализованных стран созревает всего 400-500 яйцеклеток, у женщин традиционной культуры – в племенах охотников-собирателей – менее 200 штук. Это связано с различиями в традиции деторождения: у европейских женщин рождается в среднем 1-2 ребенка, которых она кормит в среднем 3 -5 месяцев, (а известно, что лактация тормозит восстановление месячных циклов после родов), то есть у нее больший период времени остается для созревания яйцеклеток и прохождения менструальных циклов; в это же самое время у бушменов женщины рожают в среднем по 5 детей, они не делают абортов, в отличие от западных женщин, и они кормят грудью по 3-4 года, при этом овуляция тормозится, поэтому месячных циклов у них в 2 раза меньше, чем у западных женщин. Большее количество овуляторных циклов ведет к повышению риска заболевания репродуктивных органов у женщин, так как каждая овуляция связана с делением клеток, а чем больше делений – тем больше может возникнуть мутаций, ведущих к появлению злокачественных образований.
Месячные циклы у женщины регулируются изменением концентрации гормонов (верхний график на рисунке). Под действием гормонов один из покоящихся фолликулов (пузырьков) с яйцеклеткой начинает развиваться. Через несколько дней фолликул лопается и из него выходит зрелая яйцеклетка. Этот процесс называется овуляцией. Слизистая оболочка матки (эндометрий) при этом разрастается, готовясь принять оплодотворенную яйцеклетку. Если беременность не наступает, происходит дегенерация и отторжение верхнего слоя эндометрия, сопровождающееся кровотечением. Во время овуляции у женщины происходит повышение так называемой базальной температуры (то есть температуры, измеряемой ректально и вагинально сразу после пробуждения) на несколько десятых градуса (нижний график на рисунке), потом она может упасть или остаться слегка повышенной до начала менструации. У каждой женщины колебания базальной температуры индивидуальны, но более или менее постоянны при установившемся месячном цикле. Таким образом по изменению температуры можно примерно судить, когда происходит овуляция.

Ошибки при определении сроков овуляции по базальной температуре могут возникать из-за не связанных с месячным циклом изменений температуры (например, при гриппе или другом заболевании, дающем подъем температуры) или из-за сбоев цикла, которые могут возникнуть у женщины при перемене климата, стрессе или под влиянием других факторов. Пример изменения температуры в одном месячном цикле представлен на рисунке:

Яйцеклетка после выхода из фолликула сохраняет жизнеспособность примерно 24-48 часов. Спермии же после попадания в половые пути женщины жизнеспособны до 2-3 суток, далее они могут быть подвижны, но не способны к оплодотворению. Поэтому оплодотворение возможно в течение 2-3 дней до и 1-2 дней после овуляции. В остальное время зачатие произойти не может. Но на самом деле скачок температуры происходит не точно при овуляции, а при изменении концентрации гормонов, овуляцию вызывающих, поэтому точность определения дня овуляции по температурному графику составляет примерно 2 дня. Поэтому оплодотворение может произойти в 3+2=5 дней до овуляции и 2+2=4 дня после овуляции дней цикла. Осторожные люди прибавляют еще по 1-2 дня с каждой стороны. Остальные дни считаются "безопасными". Хотелось бы отметить, что цикл подчиняется эмоциональной регуляции, например, во время войны из-за тяжелой жизни, недоедания у женщин прекращались менструации, это явление называется "аменорея военного времени". Однако описаны случаи, когда муж приезжал домой с фронта на 2 дня, за эти 2 дня у женщины происходила овуляция независимо от фазы цикла, и впоследствии рождался ребенок. О том, что физиологические процессы достаточно сильно могут регулироваться нервной системой, показывает процесс родов у обезьян. У человека первые роды длятся примерно 24 часа, а у обезьян всего несколько часов, причем начинаются они обычно во время, когда стадо находится на стоянке. То есть к утру, когда стадо собирается отправляться в путь, мама готова путешествовать дальше с новорожденным. Если по каким-то причинам процесс родов к утру не завершился, а стадо уже готово идти дальше, то роды останавливаются, так как стадные животные не должны отставать от своих сородичей, и уже потом при новой остановке, роды возобновляются.
Процесс проникновения сперматозоидов в яйцеклетку называется оплодотворением. Яйцеклетка окружена несколькими оболочками, структура которых такова, что только сперматозоид собственного вида может попасть в яйцеклетку. После оплодотворения оболочки яйцеклетки меняются и другие сперматозоиды уже не могут в нее проникнуть.

У некоторых видов внутрь яйцеклетки могут проникнуть несколько сперматозоидов, но все равно в слиянии ядер участвует только один из них. При оплодотворении в яйцеклетку проникает только ядро сперматозоида, хвостик же вместе митохондриями отбрасывается, и в клетку не попадает. Поэтому митохондриальную ДНК все животные наследуют только от матери. Оплодотворенное яйцо называют зиготой (от греч. зиготос – соединенный вместе).
После оплодотворения происходит деление клетки, восстановившей диплоидный набор хромосом. первое и несколько последующих делений яйцеклетки происходят без увеличения размера клеток, поэтому процесс называется дроблением яйцеклетки.
Эмбрион (греч. "зародыш") - ранняя стадия развития живого организма от начала дробления яйцеклетки до выхода из яйца или из материнского организма (в акушерстве, в отличие от эмбриологии, термин эмбрион используют только для первых 8 недель развития, после 8-й недели называют плодом).

Эмбриогенез (эмбриональное развитие) является частью онтогенеза (индивидуального развития) – развития организма от образования зиготы до его смерти. Эмбриогенез - это процесс, в котором презумптивные зачатки занимают свои дефинитивные места.
Из школьного вы помните, что при развитии эмбриона ланцетника образуется бластула (полый клеточный шарик), из которой получается двуслойная гаструла путем инвагинации (впячивания) одной стороны бластулы внутрь.

У млекопитающих процесс происходит несколько иным образом. Дробление яйцеклетки у них приводит к образованию комочка клеток,называемого морулой. Морула подразделяется на внутреннюю часть, из которой потом развивается сам зародыш, и наружную часть, образующую полый пузырек, называемый трофобластом. Дальнейшее развитие приводит к формированию трехслойного зародыша, состоящего из внутреннего слоя – энтодермы, внешнего слоя – эктодермы, и третьего слоя между ними – мезодермы. Из каждого слоя впоследствии образуются определенные ткани и органы.
На фотографии ниже изображен червь нематода Caenorhabditis elegans (произносится как"ценорабдитис элеганс"), ближайший родственник аскариды, который интересен тем, что имеет строго фиксированное число клеток. Это дает возможность установить происхождение каждой из клеток, какая клетка из какой получилась. На рисунке показана схема происхождения разных клеток в эмбриональном развитииC.elegans.

Во время развития зародыша происходит дифференциация и перемещение его клеток с образованием тканей и органов. Рассмотрим на примере мухи-дрозофилы этот процесс. В развитии мухи происходит последовательная смена форм, значительно отличающихся друг от друга: яйцо, личинка, куколка и имаго (взрослая особь). Такое развитие называется развитием с метаморфозом (метаморфоз – греч. "изменение формы").

Цитоплазма яйцеклетки не гомогенна, в ней асимметрично распределены различные биологически активные компоненты.
У эмбриона уже определены оси тела, число и ориентация сегментов тела, из которых затем развиваются части тела взрослой мухи. Эти процесс контролируются различными наборами генов, которые называются морфогены. Они кодируют белки, которые регулируют экспрессию других генов, отвечающих за формирование органов.
Градиент концентрации белков-морфогенов определяет передне-заднюю и дорсо-вентральную (спино-брюшную) оси тела. У дрозофилы в определении дорсо-вентральной оси тела участвуют 12, главный из них ген dorsal. БелокDorsalсконцентрирован в цитоплазме на той стороне эмбриона, которая станет спинной, и проникает в ядро клеток на брюшной стороне, активируя группы генов, продукты которых необходимы для определения осей тела.
Детерминация передне-задней оси тела контролируется другими генами. Один из них называется bicoid, его мРНК сконцентрирована в цитоплазме передней части яйца (фиксирована своим 3'-концом). В результате при трансляции возникает градиент концентрации белкаBicoidот переднего к заднему концу яйца. Градиент поддерживается продолжительным периодом синтеза белка и его коротким временем жизни.Bicoid, так же как иDorsal, морфоген, он активирует гены, которые необходимы для развития головы и грудных структур. Его экспрессия ингибируется продуктом генаnanos, сконцентрированного на заднем конце эмбриона.
На следующем этапе включаются гены сегментации. Они контролируют дифференциацию эмбриона на индивидуальные сегменты. После оплодотворения транскрибируется около 25 генов сегментации, их экспрессия регулируется градиентами белков BicoidиNanos.

Гомеозисные гены . После сегментации и установления ориентации сегментов активируются так называемые геомеозисные гены. Различные их наборы активируются специфическими соотношениями концентраций белков, упоминавшихся выше.
Продукты гомеозисных генов активируют другие гены, которые определяют сегмент-специфичные особенности. Глаза в норме возникают только на головном сегменте, а ноги – только на грудных сегментах.
Гомеозисные гены кодируют регуляторные белки, связывающиеся с ДНК. Каждый из них содержит кластер нуклеотидов, называемый гомеобоксом, которые сходен во всех гомеотических генах. Он содержит 180 нуклеотидов и кодирует 60 аминокислот, функционирующих как ДНК-связывающий домен.
У дрозофилы имеется два основных кластера гомеотических генов: комплекс Antennapedia (5 генов у дрозофилы) который определяет развитие головы и передних торакальных сегментов, и комплексBithorax (3 гена) который контролирует развитие задних торакальных и брюшных сегментов. Порядок расположения генов тот же, что и сегментов, в которых они экспрессируются.
Впервые мутации гомеозисных генов были идентифицированы в 1894 году, когда Уильям Бэтсон заметил, что у растений иногда части цветка появляются на неправильных местах. Гомеозисные гены как бы определяют адрес клетки в конкретном сегменте, сообщая клеткам, в каком районе они находятся. Когда они мутируют, клетки получают "ложный адрес" и ведут себя так, будто они находятся в другом месте эмбриона
Нарушения в работе гомеозисных генов (вызванные мутациями или внешними воздействиями) нарушают формирование структур тела и могут привести, например, к образованию глаз на лапках у мухи, или к тому, что вместо антенн на голову у нее вырастут ноги (как это показано на рисунке). У человека найдены мутации в гомеозисных генах, приводящие к недоразвитию зубов, например, и к другим, более тяжелым нарушениям.

После того, как были открыты и изучены геомео-гены дрозофилы, сходные гены были найдены у всех других животных от нематоды до человека. У млекопитающих они называются Hox генами (гомеобокс-содержащими генами) и также кодируют белки, регулирующие транскрипции и определяющие структуры тела и их положение в передне-заднем направлении.

Таким образом, в эмбриональном развитии исходный градиент белков и мРНК в яйцеклетке стимулирует локальную экспрессию генов эмбриона, которая ведет дальнейшей дифференциации генной экспрессии и определяет судьбу клеток развивающегося эмбриона. Процесс, в котором формируются конечности, ткани и органы, называется морфогенезом, и определяется последовательностью переключения экспрессии групп генов, однако эти гены пока не столь детально изучены.
В процессе эмбриогенеза осуществление записанной в генах программы развития происходит в конкретных условиях среды. Взаимодействие генов и среды можно описать на следующей модели. Эмбриональное развитие можно сравнить с шариком, катящимся по наклонной поверхности с разными желобками. Такое представление эмбрионального развития, названное эпигенетическим ландшафтом, был предложен Кондратом Уоддингтоном.

Самый глубокий желобок (соответствующий наиболее вероятному пути) определяет нормальное развитие организма. Но у основного желобка есть много разветвлений, менее глубоких, соответствующих патологии, аномальному развитию организма, по ним шарик покатится с меньшей долей вероятности. Мутации меняют соотношение вероятностей разных путей (на рисунке – меняется глубина желобков), и увеличивают вероятность развития по "неправильному" патологическому пути. Однако в части случаев воздействие среды (лечение) может скомпенсировать дефект и вернуть организм на нормальный путь развития. Например, фенилкетонурия – наследственная болезнь, которую можно лечить. Суть болезни заключается в том, что у больных отсутствует фермент фенилаланингидроксилаза, превращающий аминокислоту фенилаланин в другую аминокислоту, тирозин. При блокировании нормальных путей катаболизма фенилаланина его превращение идет другими путями, обычно играющими второстепенную роль. Фенилаланин превращается в кетокислоту фенилпируват (фенилпировиноградная кислота) и другие продукты. Избыточные количества фенилпирувата легко определить по анализу мочи, и такой анализ проводится всем новорожденным в нашей стране. Одним из симптомов этой болезни является развитие умственной отсталости, которое во взрослом состоянии уже необратимо. Лечить болезнь можно в детстве специальной диетой, при которой в организме не из чего будет вырабатывать пировиноградную кислоту. Частота заболевания около 1:10 000 новорожденных, и чем раньше начато лечение – тем лучше результаты. Именно поэтому проводится тотальная диагностика новорожденных. Интересно то, что если ребенок перестает придерживаться лечебной диеты, то болезнь опять станет прогрессировать. Поэтому диету надо соблюдать до остановки физиологического роста, примерно до 20 лет, когда токсичное воздействие будет менее опасным. Больные фенилкетонурией при беременности обязательно должны соблюдать диету, так как иначе плод будет отравлен из-за нарушения обмена веществ у матери.
Таким образом, при лечении, то есть полезном воздействии внешней среды, можно вернуть развитие организма в нормальное русло. Но действие окружающей среды может быть и вредным, то есть у организма под действием внешней среды возникают отклонения развития при совершенно нормальных генах. Для примера рассмотрим один случай. В 60-х годах в Германии было сильно разрекламировано новое снотворное под названием талидомид. Среди принимавших новое лекарство были беременные женщины. Спустя некоторое время было замечено, что в стране стало рождаться много детей с патологией конечностей. У них отсутствовали длинные кости конечностей, то есть прямо от основания тела начинались кисти или ступни.
Раньше такое заболевание встречалась один раз на несколько тысяч новорожденных, и вдруг такой всплеск. Начали проводиться исследования, и выяснилось, что причина в новом лекарстве. Как оказалось, талидомид имеет большое сродство к гуанину. Взаимодействуя с ДНК, он может приводить к функциональным нарушениям. Промотор гена, отвечающего за рост и развитие длинных конечностей, содержит большое количество гуанина, таким образом, талидомид нарушает работу этих генов, и зачатки костей длинных конечностей так и не начинают развиваться. Многие из этих детей не выжили, часть из тех, кто выжил, ведут жизнь инвалидов, но есть среди них люди, которые, несмотря на инвалидность, реализовали свои возможности.
После талидомидной трагедиивсе новые лекарства проверяют на тератогенную (вызывающую нарушения развития плода) активность, и для каждого препарата указано, можно ли его принимать беременным. Однако следует учитывать, что во время беременности, особенно на ранних этапах, женщина не должна принимать лекарства, не посоветовавшись с врачом, из-за возможных вредных воздействий на плод.
В настоящее время уровень тяжелых врожденных уродств составляет 1-2%, из них около трети по генетическим причинам, около трети – из-за воздействий среды, и для трети причина неизвестна. Подбирая условия среды, соответствующие индивидуальным особенностям организма, можно скомпенсировать часть врожденных дефектов.
Взаимодействие генов и среды в процессе индивидуального развития можно сравнить с игрой в карты: хороший игрок может выиграть и с плохими картами.
|
Связь генотипа и фенотипа: варианты проявления менделевских закономерностей
|
Мы продолжим обсуждать качественные признаки, и говорить о связи генотипа и фенотипа, о вариантах проявления менделевских закономерностей, и что за этим стоит с точки зрения наших представлений о работе генетического материала.
Зачем надо изучать результаты Менделя и Моргана? Прежде всего, нам интересно узнать что-то о нас самих. Нам хочется быть богатыми, здоровыми и счастливыми (последние два признака во многом генетически контролируемые. Есть люди, которые чувствуют себя обычно счастливыми, а есть такие, которые чаще ощущают себя несчастными – эти ощущения связаны с определенными генетическими характеристиками).
Если мы знаем характер генетического контроля признака, то мы можем предсказать с определенной вероятностью фенотип на основе генотипа (если он известен). Если мы не знаем, как признак контролируется, то мы будем в полном неведении, и не сможем ничего сказать относительно признаков будущих поколений. Если мы знаем связь между генотипом и фенотипом, то мы можем сделать определенные предсказания о развитии признака (например, болезни) и, в некоторых случаях, предпринять действия полезные индивиду. Для этого нам надо установить генотип. Сейчас эта задача технически разрешима (поскольку секвенирование генома человека проведено), хотя и непомерно дорога.
В реальности нам дано лишь наблюдать проявление признака в поколениях и на основе этого создавать модель генетического контроля формирования признака, которая может быть верна лишь в данных конкретных условиях. Но, тем не менее, если мы создали такую модель, то мы можем в этом случае иметь средства для регуляции какого-то признака, в частности, повлиять возникновение или ход какой-либо болезни. Таким образом, генетический контроль и его изучение имеет большое практическое, в частности, медицинское, значение. И в основе всего лежат менделевские закономерности, которые могут проявляются по-разному в зависимости от конкретных особенностей генотипа и среды.
Мы сейчас рассмотрим, какие могут быть условия генотипа и среды, при которых эти закономерности будут выглядеть не такими, как их наблюдал Мендель, и почему это происходит. Мендель наблюдал, что при объединении задатков двух признаков в одном организме может быть проявление только одного признака. Второй задаток не проявляется. Такой тип доминирования называется полным.
Переоткрытие законов Менделя привело к выявлению других типов доминирования. Например, неполного доминирования, когда фенотип гетерозиготы является промежуточным между двумя гомозиготами. Есть еще один тип доминирования, очень популярный в последнее время в суде, – кодоминирование – в гетерозиготе проявляются фенотипы каждой из гетерозигот. Это явление имеет место, в частности, и у человека. Если у вас есть папина хромосома и мамина хромосома (а это, несомненно, так), и они различаются в миллионе позиций, что можно выявить разными методами, то это все случаи кодоминирования.
Первичным фенотипом организма является последовательность нуклеотидов молекул его ДНК.На этом фенотипе строятся все фенотипы следующих уровней. То есть при исследовании вашей ДНК проявляются все и папины, и мамины признаки; каждая из молекул ДНК проявляет свой признак независимо от присутствия другой молекулы ДНК с другим признаком: при секвенировании или при расщеплении ДНК какими-либо ферментами видны оба состояния ДНК. Кодоминантные признаки (маркеры самой молекулы ДНК) характеризуют различие между хромосомами и используются для идентификации личности или установления отцовства (число таких случаев разрешаемых в суде составляет несколько сотен в год).

Когда мы говорим о генотипе и фенотипе – это такие крайние единого процессареализации наследственной информации в индивидуальном развитии. Например, гладкая или морщинистая форма горошины, это ее фенотип. А генотип – это та специфическая последовательность нуклеотидов, которая в данных условиях определяет, что горошина будет гладкой или морщинистой. В 1999 году с менделевскими линиями гороха была проведена следующая работа. Участки хромосомы, отвечающие за форму горошины, были клонированы, секвенированы, и было установлены их особенности - различие последовательностей нуклеотидов – которые и определяли развитие гладкой или морщинистой формы горошины.
Обратите внимание, что форма горошины является конечным признаком, а формированию признака этого уровня предшествует проявление признака на многих предыдущих уровнях. Во-первых, это наличие (аллель 1) или отсутствие (аллель 2) олигосахарида, которое и приводит к той или иной форме горошины. Еще более глубоким уровнем проявления фенотипа является то, что имеется соответствующий белок (аллель 1), который необходим для синтеза олигосахарида или это тот же белок, но альтернативной структуры (аллель 2), при которой олигосахарид не образуется. Еще более глубоким признаком является РНК, с которой синтезируется этот белок. Эти РНК различны по последовательности нуклеотидов (аллели 1 и 2), что и делает различными соответствующие белки. А эти РНК различны, потому что транскрибируется с разных молекул ДНК, папиной и маминой, у которых последовательности нуклеотидов в данной позиции различны (аллели 1 и 2). Все это – проявление одного и того же фенотипа, последовательно реализуемого на каждом из уровней.
Мы имеем право говорить о фенотипе на каждом из этих многих уровней – от конкретных особенностей последовательности нуклеотидов ДНК до формы горошины. При этом, как только мы продвигаемся от ДНК выше, тем больше влияние условий окружающей среды. Например, возможность функционирования разных аллелей на уровне ДНК (транскрипция папиной и маминой копии гена) будет мало зависеть от температуры, а возможность функционирования тех же аллелей на уровне белка может критически зависеть от температуры. При некоторых температурах белок (например, аллель 1) будет работать, а другой (аллель 2) не будет работать. Как только мы выдвигаемся на более высокий уровень в реализации фенотипа, появляется больше возможностей для влияния окружающей среды на проявление признака.
И наоборот, чем ближе мы продвигаемся к генотипу, тем предсказуемее связь между генотипом и фенотипом. При идентификации человека в судебно-медицинской экспертизе чаще всего используются маркеры, которые различают одни и те же участки гомологичных (папины и мамины) хромосом. Обычно эти различия - микровставки в данном месте молекулы ДНК. Принцип метода таков. Мы можем размножить in vitro фрагмент молекулы ДНК в нужном месте хромосомы, например, с координатами от позиции N до позиции N+100. Этот фрагмент мы выбрали потому, что в нем у каждого человека в каждой хромосоме есть вставка, например, динуклеотид СА. В каждой хромосоме длина этой вставки может быть своя. Допустим, в одной хромосоме этот динуклеотид повторен в данном месте 10 раз, а в том же месте другой хромосомы – 15 раз. При размножении этого участка хромосомы мы получим фрагменты двух длин 100 +2*10=120 пар нуклеотидов и 100+2*15=130 пар нуклеотидов. Фрагменты каждого из этих размеров отличимы после их фракционирования электрофорезом в геле.
Пусть и папа и мама гетерозиготны по этому локусу, подобно гибридам первого поколения Менделевского моногибридного скрещивания. Обозначим их генотипы по характеристической длине размножаемых фрагментов ДНК – 120/130. Тогда их дети будут иметь такие генотипы 1 120/120 (гомозиготы по аллелю 120) : 2 120/130 (гетерозиготы) : 1 130/130 (гомозиготы по аллелю 130). Естественно, присутствие в образце фрагмента ДНК одной длины не влияет на возможность детекции фрагмента ДНК другой длины, то есть, используемые маркеры ко-доминантны. Они обе проявляются. Понятно, что здесь проявляются те же менделевские закономерности передачи и расщепления признаков, хотя в качестве признака мы использовали морфологию самой молекулы ДНК – различную длину фрагмента молекулы в данном месте хромосомы.
После переоткрытия законов Менделя выяснялось, что Мендель то в одном как бы как бы не прав, то в другом как бы не прав. Обнаруживались дополнительные обстоятельства, которые модифицировали проявление менделевских закономерностей.
Например, число классов генотипов и фенотипов зависит от плоидности, даже если аллелей всего два. К примеру, эндосперм в зерне кукурузы триплоидный. У него два генома маминых, а один папин (из пыльцевого зерна). Получали растения кукурузы, которые различались по числу доминантного аллеля гена Y. Этот аллель необходим для образования витамина А. Когда в эндосперме была одна копия аллеля Y, то Если относительное количество витамина А при одной копии аллеля Y составляло единицу, то при двух копиях – 2,2, а при трех копиях – 3,3. То есть выраженность конкретного признака может зависеть от дозы гена. А может и не зависеть – как в случае полного доминирования одного аллеля над другим. Все зависит от конкретного механизма формирования признака.
То, что от дозы гена зависит количество его продукта, используется в биотехнологиях, как основа всех современных процессов, основанных на рекомбинантных ДНК. Во всех этих случаях в клетке увеличивают дозу гена. Увеличение дозы гена приводит к нужным нам результатам, но клетка от этого может умереть. Обычно при производстве это безразлично. Критерием эффективности в биотехнологии является получение целевого вещества с минимальными затратами. Впрочем, если бы могли получить корову, которая бы дала сразу цистерну молока, а потом бы умерла, это было бы очень технологично, но не этично. Однако с бактериями так и поступают: выращивают бактерии в идеальных условиях, а затем включают синтез, который стоит бактериям жизни.

Например, с помощью технологий рекомбинантных ДНК сконструированы бактерии, превращающие сахар в аминокислоту треонин. Более половины углерода, поглощенного такими бактериями превращается в единственный целевой продукт. Меньше половины сахара превращается в тысячи других углерод-содержащих соединений клетки, а, кроме того, расходуется еще и как источник энергии, чтобы все это синтезировать и просто жить. Треонин накапливается в среде до концентрации в 100 грамм на литр, что близко к пределу его растворимости. Мировые объемы производства рекомбинантных продуктов составляют десятки тысяч тонн в год (треонин) и даже сотни тысяч тонн (лизин).
В менделевском случае каждый признак формировался под действием как бы одного гена, который на другие признаки не влиял (ведь растение с гладкими семенами могло быть разной высоты, иметь разного цвета семена и цветы и др.), На самом деле формирование каждого признака требует работы многих генов, и большинство генов влияют более, чем на один признак. Например, относительное содержание каждого из трех метаболитов в эндосперме кукурузы (на слайде) зависит от генотипа по каждому из трех исследованных генов.

Видно, что на каждый признак влияет каждый из трех генов, и таких генов наверняка больше. Для формирования каждого признака нужно много генов. Объясним это на примере слуха. Чтобы человек слышал, нужны многие структуры уха: молоточки, наковаленки, барабанные перепонки, и если не будет какого-то белка, нужного, чтобы сформировать эти структуры, то человек не будет слышать. Поэтому глухота у человека может возникать из-за дефектов в десятках известных генов (контролирующих много элементов слухового аппарата, передачу сигнала в нервной ткани т.д.). При этом разные генотипы могут приводить к одному фенотипу (глухота). Внешне одинаковые фенотипы, обусловленные разными генетическими причинами, называются фенокопиями. Мутации глухоты редки и обычно рецессивны. Глухота проявляется обычно как моногенный аутосомный дефект с наследованием по менделевскому типу. Известны случаи, когда муж и жена, оба глухие, хотели бы, чтобы и их будущий ребенок был глухим, чтобы он также принадлежал их сообществу. Этот пример показывает – понятие нормы и болезни относительно. Оно зависит, в том числе, и от отношения индивида к своей особенности и от отношения общества к этой особенности.
Менделевские закономерности при взаимодействии генов (на примере глухоты). Допустим, что глухота определяется рецессивными мутациями в 10 разных генах и частота этих мутаций в популяции одинакова.
Вопрос 1.Какова вероятность того, что у двух глухих родителей будет глухой ребенок? Решение. Каждый из родителей - гомозигота по мутации в одном из генов, нужных для развития слуха. Вероятность того, что второй родитель несет мутацию в том же гене, что и первый составляет 1/10. Следовательно, вероятность того, что ребенок от такого брака будет глухим, составляет 10%, а того, что он будет слышать – 90%. Запись генотипов будет при этом следующая: родителиааВВхААbb, потомство первого поколенияАаBb.
Вопрос 2 . Какова будет доля слышащих потомков от скрещивания индивидов первого поколения? (Чтобы не смущать друг друга будем считать, что этот опыт проводится, конечно, не на людях, а на мышах – основном модельном генетическом объекте из млекопитающих). Решение. Расщепление по каждой паре аллелей происходит независимо друг от друга. То есть скрещивание АаBbхAaBbможно записать как два независимых скрещивания. По гену А: родителиАахАа. Расщепление у потомков по фенотипу составит 3 : 1 (3 слышащих к 1 глухому). При этом у всех слышащих есть хотя бы один доминантный аллельА. Такой генотип записывается в формеА- . Генотип глухих потомков –аа(гомозиготы по рецессивному аллелю). Тогда генотипы соответствующие слышащим и неслышащим потомкам скрещивания можно представить как 3А- : 1аа.
Таким же образом запись потомков по генотипу при расщеплении аллелей по локусу Вможно представить как 3В- : 1bb.
Поскольку расщепление по аллелям одного локуса происходит независимо от расщепления по аллелям другого локуса, то генотип потомков при расщеплении по двум локусам одновременно можно записать в следующем виде:
(3А- : 1аа) х (3В- : 1bb) = 9A-B- : 3aaB- : 3A-bb: 1aabb
Чтобы индивид слышал, ему необходимо иметь хотя бы один аллель дикого типа по каждому из локусов. Таких индивидов 9 (генотипы А-В-). Чтобы индивид не слышал, достаточно гомозиготности по любому из двух рецессивных генов. Таких индивидов 7 (генотипыaaB-,A-bbиaabb)
Таким образом, при скрещивании дигетерозигот соотношение слышащих потомков к неслышащим составит 9:7.
Отдельный ген может влиять на несколько признаков, в развитие которых он вовлечен.Такой эффект действия гена называется плейотропным (множественным). Например, альбинизм вызывается дефектом одного гена, контролирующего образование меланина. При этом меняются одновременно такие признаки как цвет волос (белый), цвет кожи (белый) и цвет глаз, (он будет красным от цвета кровеносных сосудов, лежащих за радужной оболочкой глаза). Множество признаков определяется геном SRY, контролирующим формирование всех признаков мужского пола у млекопитающих. Перенос генаSRYиз Y-хромосомы самца в геном самки мыши, приводил к развитию особи с огромным количеством внешне наблюдаемых признаков мужского пола.
У Менделя получились одинаковые численные соотношения при расщеплении аллелей многих пар признаков. Это в частности подразумевало одинаковую выживаемость индивидов всех генотипов, но это может быть и не так. Бывает, что гомозигота по какому-нибудь признаку не выживает. Например, желтая окраска у мышей может быть обусловлена гетерозиготностью по одной из пар аллелей (слайд). При скрещивании таких гетерозигот друг с другом следовало бы ожидать расщепление по этому признаку соотношении 3:1. Однако, наблюдается расщепление 2:1, то есть 2 желтых к 1 белой (рецессивная гомозигота). Показано, что доминантная (по окраске) гомозигота не выживает уже на эмбриональной стадии. Этот аллель одновременно является рецессивной леталью (то есть рецессивной мутацией, приводящей к гибели организма).

На следующей картинке показаны типы групп крови, их наследуемость. Этот пример приведен для того, чтобы сказать, что у одного гена может быть не только два аллеля. У Менделя их было всего два для каждого признака: например желтая и зеленая окраски, третьего не дано. Но на самом деле аллелей по каждому локусу может быть много. Группы крови различаются по тому, какой тип антигена мы вырабатываем. Может вырабатываться антиген типа АилиВ, но если одна хромосома ответственна за типА, а другая за типВ, то будут вырабатываться оба типа антигена. Перед нами случай кодоминирования на уровне белка, когда наличие одного типа антигена не препятствует наличию другого. Четвертый тип – когда антигены вообще не вырабатываются. Какое практическое значение это имеет для нас? Это были первые генетические исследования, проведенные во время первой мировой войны в австрийской армии, так как во время войны всегда была острая необходимость переливания крови, которое не всегда оказывалось удачным, то есть человек от переливания крови иногда умирал.

Переливать кровь наиболее безопасно между людьми, у которых одинаковая группа крови. Антигены групп крови у них одинаковые, антител против них нет и агглютинация (слипание клеток крови) не произойдет.
Если донором (то есть сдающим кровь) является человек, у которого ни одного из антигенов нет (группа крови 0), то его кровь можно переливать кому угодно – она не будет вызывать агглютинации у реципиентов (тех, кому кровь переливают) с любым типом крови. Таким образом, люди с типом крови 0 являются универсальными донорами. И наоборот, в крови людей с группой АВ не содержатся антитела ни на один из этих антигенов (ни на А, ни на В) и эти люди счастливы быть универсальными реципиентами – переливать кровь им можно от людей своей и всех остальных групп крови. То есть человек с группой крови АВ может принять кровь с любым типом. Самый плохой случай для реципиента – нулевая группа крови, которая будет реагировать с группами крови А, В и АВ. Два разных гена могут находиться в разных локусах и влиять на один и тот же признак. Например, у дрозофилы могут быть разный цвет глаз, от коричневого (рецессивная гомозигота по одному гену), через разные оттенки красного к белому (рецессивная гомозигота по другому гену). Красные глаза имеет муха дикого типа (например, гомозигота по всем аллелям дикого типа), а также гетерозигота по двум генам, каждый из которых нужен для пигментации глаза (см. рис. слева). Взаимодействие двух генов в таком случае называют комплементарностью (дополнительностью). При скрещивании дигетерозигот дает расщепление по фенотипу в отношении 9:3:3:1. Из четырех фенотипических классов по окраске глаза самый многочисленный – 9, это окраска дикого типа, а самый малочисленный – 1, отсутствие окраски из за дефекта обоих генов, определяющих образование разных пигментов.

Такая же ситуация у человека с окраской волос, у нас есть разные типы меланина: эу- и фикомеланин, которые в разном сочетании дают разные оттенки, например рыжий.Справа на рисунке приведен тот же пример, только на растениях. Гены в различных локусах могут давать вклад в развитие одной и той же фенотипической характеристики: у перца 2 разных гена нужны для образования двух разных пигментов, что приводит к расщеплению по типу дигибридного скрещивания. Это используется при получении определенного цвета перца. Чтобы получит потомство с нужным цветом плода необходимо знать генотипы родителей и правильно их подбирать для скрещивания.

На животных это применяется на пушных зверьках в звероводческих фермах. За счет определенного подбора аллелей десятков известных генов, определяющих окраску шерсти, можно получить большое разнообразие окрасок шкур пушных зверьков, причем заранее планировать сколько и каких типов окраски получится.
Если два разных гена взаимодействуют при формировании признака и их действие складывается, то такое действие генов называют кумулятивным (накапливающимся). Например, степень красной окраски зерна пшеницы определяется двумя разными генами, в каждом из которых есть неполностью доминантный аллель А.
Таким образом, наблюдается много вариантов численных соотношений классов в потомстве дигибридного скрещивания в зависимости от типа взаимодействия генов. Если взаимодействия нет, то отношение будет 9:3:3:1. Может быть так, что появление в генотипе рецессивного гена аведет к тому, что генВв фенотипе не проявляется. Тогда соотношение будет 9:3:4. То же, но с доминантным геном – будет 12:3:1. И т.д. То есть это показывает, что Мендель был прав только в достаточно частном случае, когда гены не влияют друг на друга, но может быть еще много других вариантов.

Породы собак: для того, чтобы получить какие-то определенные зафиксированные признаки породы, проводили близкородственное скрещивание, пока эти признаки не закреплялись в гомозиготном состоянии, и далее при скрещивании собак одной породы расщепление по этому признаку больше не может происходить – признак становится стабильным, типичным для породы.

Каждая порода собак обладает своими признаками, и эти признаки стабильны в поколениях, поскольку каждую породу характеризует свой набор гомозиготных локусов, определяющих эти признаки. У каждой породы собак как свой набор гомозиготных локусов (у других пород они могут быть и гетерозиготны). Однако у одних пород данный локус гомозиготен по одним аллелям, а других тот же локус может быть гомозиготен по другим аллелям, что и определяет стабильные различия между породами (например, по окрасу). Ясно, что у каждой породы какие-то локусы остаются гетерозиготными, и собаки одной породы по этим признакам будут различаться, но это те признаки, которые данную породу не определяют.

Если скрестить собак разных пород, то получится дворняжка. У нее по большинству генов гетерозиготное состояние, соответствующее предковым формам. Дарвин в свое время подметил, что если скрещивать разные породы голубей, получается все время сизарь. Это происходит из-за утраты у гибрида гомозиготности характерной для каждой из родительских пород. А в гетерозиготных локусах один из аллелей, как правило, оказывается дикого типа, поэтому и выглядят гибриды сизарями – это дикий тип для всех пород голубей.
По генам окраски меха у норки в настоящее время установлено около 20 сериймножественных аллелей. При скрещивании гетерозиготных норок между собой во втором поколении будет расщепление, типичное для дигибридного скрещивания в соотношении 9:3:3:1, или 56% норок с дикой окраской меха, 19% – алеутских, 19% – платиновых и 6% с сапфировой окраской меха. Вавилов сформулировал закон гомологических рядов: если у какого-то вида есть какой–то набор вариантов признаков, и мы рассматриваем близкородственный вид, то у него тоже может быть выявлен тот же набор признаков. Этот закон основан на том, что у близкородственных видов и набор аллелей в ДНК может быть сходен.

Гены могут находиться не только в ядре.Гены могут наследоваться однородительски, например, по материнской линии: окраска листа определяется генами хлоропластов, а не ядра, и наследуется только от матери. То есть, какая бы ни была мужская особь, потомство будет все однотипно – все в мать и только в нее.
У человека тоже есть признаки, которые определяются митохондриями, то есть наследуются по материнской линии. Митохондрии находятся в цитоплазме клетки, которая достается потомкам только от матери. К мутациям и болезням связанным с митохондриями относятся, например, дефекты зрения и дефекты энергетического обмена.
Часть ядерных генов также наследуется только однородительски. Это гены Y-хромосомы, которая наследуется только от отца к сыну. Те гены в Y-хромосоме, которые не имеют пары в Х-хромосоме, наследуются однородительски – от отца к сыну. У человека один из таких генов определяет признак «волосатые уши» – у женщин такой признак не проявляется – гена этого нет, нет и его аллеля, из-за которого уши волосатые.
Проявление признака может зависеть также от пола индивида. Существуют признаки, гены которых могут находиться в аутосомах или половых хромосомах обоих полов, но проявляются лишь у одного из них. Такие признаки называют ограниченными полом.Например, быки несут гены, определяющие молочность дочерей, но гены свое действие у быков не проявляют.Гены казеинов – белков молока– это случай наиболее строгого контроля транскрипции ограниченной полом. Они действуют у особей как женского, так и мужского пола. Разница уровней экспрессии между ними составляет 1 на 100 000. Петухи также несут гены яйценоскости и размера яиц дочерей, хотя у петухов действие этих генов подавлено. Когда проводится селекция, учитывается этот факт. Здесь существенен генетический контроль признаков.
Как проявление действия гена, так и тип доминирования может зависеть от пола. Плешивость у человека наследуется аутосомно и моногенно. Аллель плешивости у мужчин доминирует, а у женщин рецессивен. У мужчин для облысения достаточно присутствия одного аллеля плешивости. При этом частота аллеля равна частоте встречаемости фенотипа. У женщин для облысения необходимо присутствие обоих аллелей плешивости. Экспрессия некоторых генотипов может зависеть от внешних условий. Ниже показаны два кролика, один из которых с темными пятнами. Аллель гималайской окраски у кролика температурочувствителен. При повышенной температуре белок не функционален и необходимый пигмент не образуется, а при нормальной температуре получается кролик, у которого некоторые участки шкуры окрашены.

Мутанты, проявление которых зависит от условий среды, сыграли очень важную роль в биологии вообще и в молекулярной биологии в частности, в понимании того, как работает генетический материал. Многие генетические феномены, были открыты и исследованы после получения и использования условно летальных мутантов (условно смертельные). Наиболее распространенные мутанты такого типа – температурочувствительные. Получены мутанты многих микроорганизмов и вирусов, которые не способны жить при повышенной температуре (ts-мутанты). У такого мутанта структура белка при повышенной температуре изменена, и белок не функционален (леталь), а при нормальной температуре структура обычная, и белок функционален. При этом возможно исследовать особенности функционирования белка и его роли в формировании какой-либо структуры или сложной функции. Для этого клетки растят до нужной стадии, в условиях, когда белок функционален, затем разделяют клетки (обычно суспензию) на две части, в одной температура остается нормальной, а в другой –поднимается, так что белок теряет функциональность.
Есть также другой тип условно летальных мутаций– нонсенс-мутации, летальные на одном генетическом фоне и нелетальные на другом. Нонсенс-мутация приводит к изменению кодона для какой-либо аминокислоты в данном белке на стоп-кодон трансляции. Белок при этом образуется укороченный и нефункциональный. Нонсенс-мутация подавляется (супрессируется), в тех клетках, где образуется супрессорная тРНК. Супрессорная тРНК, это измененная тРНК, у которой антикодон по-прежнему узнает стоп-кодон в матричной РНК, но вместо терминации (окончания) трансляции белка на этом участке происходит подстановка определенного аминокислотного остатка. Белок с нонсенс-мутацией синтезируется полноразмерным и функциональным в присутствии тРНК.
При создании модели наследования признакадля выяснения его генетического контроля начинают с проверки типа наследования по базовой, простейшей схеме - менделевской и, если соответствия нет, то добавляют к этой схеме дополнительные условия, понятные на сегодня:
неравную вероятность образования и сочетания гамет и выживания потомства;
однородительский тип наследования (Y – волосатые уши, мтДНК - болезнь Лебера)
взаимодействие аллелей: наличие доминирования;
взаимодействие генов;
сцепление генов;
ограничения признака полом;
вероятность проявления признака при данном генотипе;
влияние среды (не всегда формализуемое).
Напомним, что менделевский характер наследованияотносится к признакамкачественным.Сколько Меделирующих признаков известно у человека? Они зафиксированы в базе данных OMIM («Менделевское наследование у человека»). Их там перечислено около пяти тысяч признаков. А мы у себя можем насчитать гораздо больше признаков. Огромное количество остальных наших признаков зависит от действия многих генов. Здесь следует отметить важный идеологический аспект. Общей модели, по которой происходит формирование признаков под воздействием многих генов, сейчас нет. Наверное, ее и быть не может. Чтобы понять, как данный фенотип возникает на фоне данного генотипа, нам нужно еще узнать, как конкретно реализуется каждый этап передачи генетической информации: что происходит с модификацией ДНК, с какой из многих возможных вариантов РНК образуется с данного участка генетической информации, как модифицируется белок, образуемый с этой мРНК. В ДНК заложена лишь возможность всех этих вариантов реализации генетической информации. Произойдет ли это или нет, зависит от среды. Понятие среды мы пока толком формализовать не можем. Но если, подобно структуре ДНК, мы их будем знать структуру и модификацию РНК и белков, то понимание связи генотипа с фенотипом будет гораздо более глубоким, а предсказательная сила наших знаний будет больше. Как бы там ни было, одного знания генотипа часто недостаточно для предсказания фенотипа. Поэтому мы должны изучать следующие этапы экспрессии генетической информации.
В заключительной части лекции речь пойдет об определении пола и признаках, сцепленных с полом. Ниже показано, как выглядят половые хромосомы человека. Х-хромосома по длине ДНК больше чем Y-хромосома приблизительно в три раза.

А так выглядят клетки, транспортирующие эти хромосомы. Маленькая образована организмом генотипа XY, а большая – генотипаХХ. Такое соотношение физических размеров имеют половые клетки.
Так выглядит человек после того, как он уже начал развиваться:

Это игла, и на ее конце находится несколько поделившихся клеток, полученных из оплодотворенной яйцеклетки. Такой наш размер на стадии 4-5-ого деления.
Еще раз напомню, что у нас цикл развития двухстадийный. На нижней части рисунка изображена диплоидная стадия, период нашей долгой и счастливой жизни, которая с точки зрения генетического процесса заканчивается мейозом, после которого мы начинаем жить как бы в следующей стадии, гаплоидной. Это гаметы, у нас они не обладают способностью к самостоятельному существованию, хотя у многих растений именно гаплоид является основной частью жизненного цикла. Существование гамет заканчивается оплодотворением и появлением опять диплоидного организма.

Х и Y хромосомы сильно отличаются. Они гомологичны только в так называемых псевдоаутосомных зонах, которые присутствуют и в папиных и в маминых хромосомах, как и обычные аутосомы. Спермии могут быть двух типов: Х и Y, а яйцеклетки только одного типа – Х и Х. Поэтому расщепление получается 1:1 (две женщины и два мужчины). Именно расщепление 1:1 по полу на первом этапе исследования признака, сцепленного с полом, – окраски глаз у дрозофилы – привело Моргана к мысли о том, что гены находятся в хромосомах. Пол определяется хромосомами и гены находятся именно в них.

Не у всех организмов, в отличие от человека, пол однозначно определяется при слиянии гамет. Ниже показан моллюск, у которого в нижней части женские клетки, а в верхней части только мужские. У некоторых пресмыкающихся мужской пол не формируется при повышенной температуре (в частности, у крокодилов), образуются только женские организмы.

У дрозофилы всего четыре пары хромосом, три пары хромосом называются аутосомы и одна пара половых хромосом. У самок набор хромосом ХХ, а у самцаХY– как и у человека (у самки на конце брюшка находится яйцеклад, поэтому у нее брюшко заостренное, а у самца оно овальное, - так их отличают).

У плодовой мушки пол определяется не столько наличием той или иной половой хромосомы, а отношением числа Х-хромосом к числу гаплоидных наборов аутосом (А). Это видно на схеме ниже. НаборХХ+2А(2 Х-хромосомы и 2 набора аутосом) дает соотношение 1,0 – такое соотношение является нормой для самок. НаборХY+2А(1 Х-хромосома и 2 набора аутосом) дает соотношение 0,5 – такое соотношение является нормой для мужского пола. Если оказывается, что Y-хромосома утеряна (набор 1Х+2А, соотношение 0,5), то образуется мужской организм, хоть Y-хромосомы и нет. А если на фоне двух Х-хромосом добавилась Y-хромосома (набор 2Х+2А), пол формируется женский, хоть Y-хромосома и присутствует. У человека, сколько бы ни было Х-хромосом, если есть хоть одна Y-хромосома, то фенотип будет мужской.
Таким образом, у человека и мухи, при внешне одинаковой хромосомной формуле нормы (гомогаметный пол, ХХ, женский; гетерогаметный пол,ХY– мужской), механизм генетического контроля образования пола различен.
У птиц и ряда других групп организмов гомогаметный пол (ХХ) – мужской, а гетерогамтный пол (XY) – женский, что противоположно тому типу хромосомного определения пола, который оказался зафиксирован эволюцией в линии человека и других млекопитающих.

На следующем слайде показано месторасположение SRY-гена, отвечающего за формирование признаков мужского пола. Уже говорилось, что если в геном мыши, которая должна развиться самкой, пересадитьSRY-ген, то у такого организма разовьется мужской фенотип. Человек почти не отличается от мыши в этом плане. У человека известны случаи, когда индивид кариотипически (то есть по типу хромосом в ядре) соответствовал женщине (в частности с двумя ХХ хромосомами и без наблюдаемой Y-хромосомы), но проявлял фенотип мужчины. В таких индивидов была обнаружена хромосомная перестройка – генSRYбыл транслоцирован (перемещен) на одну из аутосом.

На внизу на рисунке представлен случай отклонения по составу половых хромосом – синдром Тернера: индивид генотипа Х0(Y-хромосомы нет, а Х-хромосома одна). Фенотип индивида - женский. Отклонений в поведении нет, хотя при большем дисбалансе в генотипе наблюдаются проблемы в умственном развитии.
Другой случай отклонения по составу половых хромосом - это синдром Клайнфельтера: генотип XXY, фенотип мужской. Хотя две Х-хромосомы присутствуют, как в норме у женщин. То есть для определения пола у человека важно, чтобы присутствовала Y-хромосома.

Напомним, что гены, локализованные в Y-хромосоме и не имеющие пары в X-хромосоме, наследуются только от отца к сыну. У человека аллель одного из таких генов определяет признак «волосатые уши». Через мать к ребенку могут передаваться гены, находящиеся в митохондриях, но т.к. генов в митохондриях на три порядка меньше чем в ядре, то и отклонений, передающихся только материнской линии лишь тысячная часть от всех известных для человека.
Избыток или недостаток какой-то хромосомы в геноме приводит к очень серьезным нарушениям у человека. Для примера можно привести трисомию по 21-ой хромосоме – синдром Дауна. Причина заболеваний при утрате или избытке любой хромосомы - дисбаланс в количестве продуктов генов этой хромосомы относительно количества продуктов генов других хромосом(все – по две). Дисбаланс по всем крупным хромосомам человека вообще не совместим с жизнью.
Х хромосома одна из крупных хромосом человека. У женщин таких хромосом две, а у мужчин – одна. Но оба пола живы и здоровы – хоть на себя погляди. Почему разница в числе копий Х-хромосомы не смертельна, в отличие от ситуации с другими хромосомами?
Чтобы сбалансировать действие генов Х хромосомы у мужчин (где Х-хромосома одна) и у женщин (где Х-хромосомы две) природа предусмотрела инактивацию одной из Х-хромосом в каждой клетке женского организма. У женщин одна Х-хромосома инактивирована, то есть гены в ней не работают, а работают только в другой хромосоме – не инактивированной. Инактивиация вызвана компактизацией хромосомы даже в интерфазе. Поэтому компактизированная хромосома видна при окрашивании – в виде так называемого тельца Барра (картинка). Это один из диагностических признаков женского пола у млекопитающих, что у человека, что у слона. Некомпактизованная Х-хромосома, как и все аутосомы, не окрашивается и не видна в интерфазе.
На следующем рисунке показана кошка черепаховой окраски. Поскольку это кошка, а не кот, то генотип у нее ХХ. В каждой клетке женского организма одна из Х-хромосом инактивирована (ее видно как тельце Барра). На Х-хромосоме у кошки находится один из генов, определяющих окраску шерсти. У этого гена известны разные аллели, то есть определяющие разный цвет шерсти. Кошка на рисунке гетерозиготна по этим аллелям. Одна из Х-хромосом в каждой клетке инактивирована, поэтому только один из двух цветов может проявится в волоске, сформированном из данной клетки. Поэтому одни волоски одного цвета, а другие – другого. Какая из Х-хромосом инактивируется (папина или мамина), и, стало быть, аллель какого цвета будет работать - дело случая. Поэтому такая кошка и имеет черепаховую окраску. Кстати, сыновья у нее будут двух разных цветов в соотношении 1:1.

Еще одно доказательство того, что гены находятся в хромосомах, было получено на мухах дрозофилах, называемых латеральными (боковыми, «сторонними») гинандроморфами. У таких мух после первого деления яйцеклетки в одной из клеток была утрачена Х-хромосома. Эта клетка при последующих делениях дала клетки с генотипомХ0, поэтому половина тела таких мух сформирована как у самца и во ее всех клетках (на рисунке – правая половина) цитологически наблюдается утрата одной из Х-хромосом. Левая половина тела, происходящая из клеток с нормальным генотипомХХ, формируется как самка.

В представленном на рисунке случае муха была гетерозиготна по гену окраски глаза и по гену, влияющему на форму крыла. Эти гены расположены в Х-хромосоме. Поэтому правый глаз мухи неокрашен: проявилась рецессивная мутация white, присутствующая в одной из Х-хромосом. Крыло изменено по форме. Левый глаз окрашен и крыло нормальной формы, поскольку в нем остались обе Х-хромосомы, в одной из которых присутствует рецессивный генwhite, а в другой – доминантный генw+.
|
|

Вопросы для самопроверки к темам:№2 и №3
Охарактеризуйте строение прокариотов, расскажите их значение в природе жизни человека.
Сравните способы питания автотрофных и гетеротрофных организмов.
Докажите, что обмен веществ и энергии являются основной жизнедеятельностью клетки.
Дайте сравнительную характеристику аэробным и анаэробным организмам. Объясните их роль в природе.
Раскройте суть процесса фотосинтеза и его планетарное значение.
Приведите примеры усложнений организации в животном царстве и объясните их значение.
Дайте сравнительную характеристику процессов митоза и мейоза.
Как происходит процесс оплодотворения у животных? Какое значение имеет искусственное оплодотворение в сельском хозяйстве.
Охарактеризуйте основные периоды онтогенеза многоклеточных организмов
Объясните значение закона гомологических рядов наследственной изменчивости.
Раскройте суть хромосомной теории наследственности.
Охарактеризуйте гены и хромосомы, как материальные носители наследственности.
Раскройте роль генотипа и среды в повышении производительности сельскохозяйственных растений и животных.
Охарактеризуйте методы изучения наследствнности человека. Приведите примеры наследственных болезней человека.
Обоснуйте значение наследственной изменчивости в эволюции органического мира.
Объясните значение модификационной изменчивости в жизни организмов.
Раскройте суть законов наследственности при моногибридном скрещивании.
Раскройте суть закона наследственности, установленного Г. Менделем при дигибридном скрещивании.
В виду ограниченийпо объему страниц учебно-методического комплекса по биологии темы лекционного курса : №4 Основы анатомии высших растений и морфология вегетативных органов, №5 Основы анатомии и физиологии животных ,№6 Разнообразие форм жизни достаточно хорошо представлены в прилагаемой к УМК «Биология» полного лекционного курса в электронном виде. Контрольные вопросы по всему курсу, в том числе к темам № 4-6 представлены в конце учебного комплекса.