

2.5. Лесные биогеоценозы на суглинистых почвах
(рамени)
Рамени — старинное слово, обозначающее еловые, темнохвой-ные леса. Коренные ельники Русской равнины образуют несколь-ко экологических групп, занимающих разные экотопы в пределах моренных равнин и конечно-моренных возвышенностей с преоб-ладанием моренных валунных суглинков по всей верхней толще. Соответственно верхние корнеобитаемые горизонты почв в ко-ренных ельниках по механическому составу относятся обычно к легким суглинкам, реже к средним суглинкам или глинам.
Представление о «сосновых» и «еловых» почвах бытовало в на-роде еще с давних времен, но лишь в XX в. в связи с развитием геохимии ландшафта были выяснены действительные причины почвенных предпочтений сосны и ели. Выяснилось, что ель фор-
мирует чистые древостой, вытесняя сосну лишь при достаточно благоприятном режиме кальция в почве, причем не только на дренированных, но и на переувлажненных почвах с подтоком вод, обогащенных карбонатами кальция и магния.
По этой же причине ель совсем неплохо себя чувствует и на тяжело супесчаных почвах конечных морен, обогащенных глини-стыми частицами и рухляком обломочных пород. Однако и сосна при прочих благоприятных условиях может заходить на суглини-стые почвы. Мы отмечаем это обстоятельство отнюдь не затем, чтобы окончательно запутать начинающих ландшафтоведов, а лишь для того, чтобы подчеркнуть: в природе нет твердых и жестких зависимостей между любыми двумя парами факторов (например, в паре механический состав почв — доминанта лесного биогеоце-ноза). Именно этим и объясняется отсутствие резких переходов и границ между растительными сообществами — во-первых, и чрез-вычайное разнообразие лесных ландшафтов — во-вторых.
Кроме того, распространению еловых лесов во многом сопут-ствует неглубокое залегание карбонатной морены, которая очень широко распространена в центральной и восточной частях юж-ной тайги.
Хотя в целом для раменевого экологического ряда в пределах лесной зоны характерны суглинистые почвы, строение почвен-ного профиля может быть разнообразным. В центральной части южной тайги чаще встречаются двучленные почвы с верхней лег-косуглинистой или супесчаной толщей, подстилаемой на глубине 50 — 80 см обычно моренным валунным суглинком. Глубина зале-гания тяжелого суглинка определяет «шансы» сосны участвовать в составе еловых древостоев.
В зону произрастания ельников попадают и озерно-леднико-вые равнины, и надпойменные террасы, и широкие озерные низ-менности, поскольку в пределах всех этих поверхностей мы также сталкиваемся с залеганием более легких по механическому составу верхних слоев на расположенных глубже суглинках — озерных, аллювиальных и т.д.
В силу экологических особенностей ели раменевая группа лише-на «сухого» местообитания, который был бы аналогичен лишай-никовым борам, и начинается сразу с центрального члена ряда, представляющего экологическую «норму» — ельника-кисличника. Кислица очень теневынослива, и поэтому она одна способна фор-мировать негустой покров по затененным местам в сомкнутых груп-пах деревьев и подроста или под кронами крупных елей.
Ельники-кисличники (рис. 2.13) развиваются на своеобразных слабоподзолистых суглинистых почвах, с нечетким разделением элювиального горизонта (А) и горизонта вмывания (В).
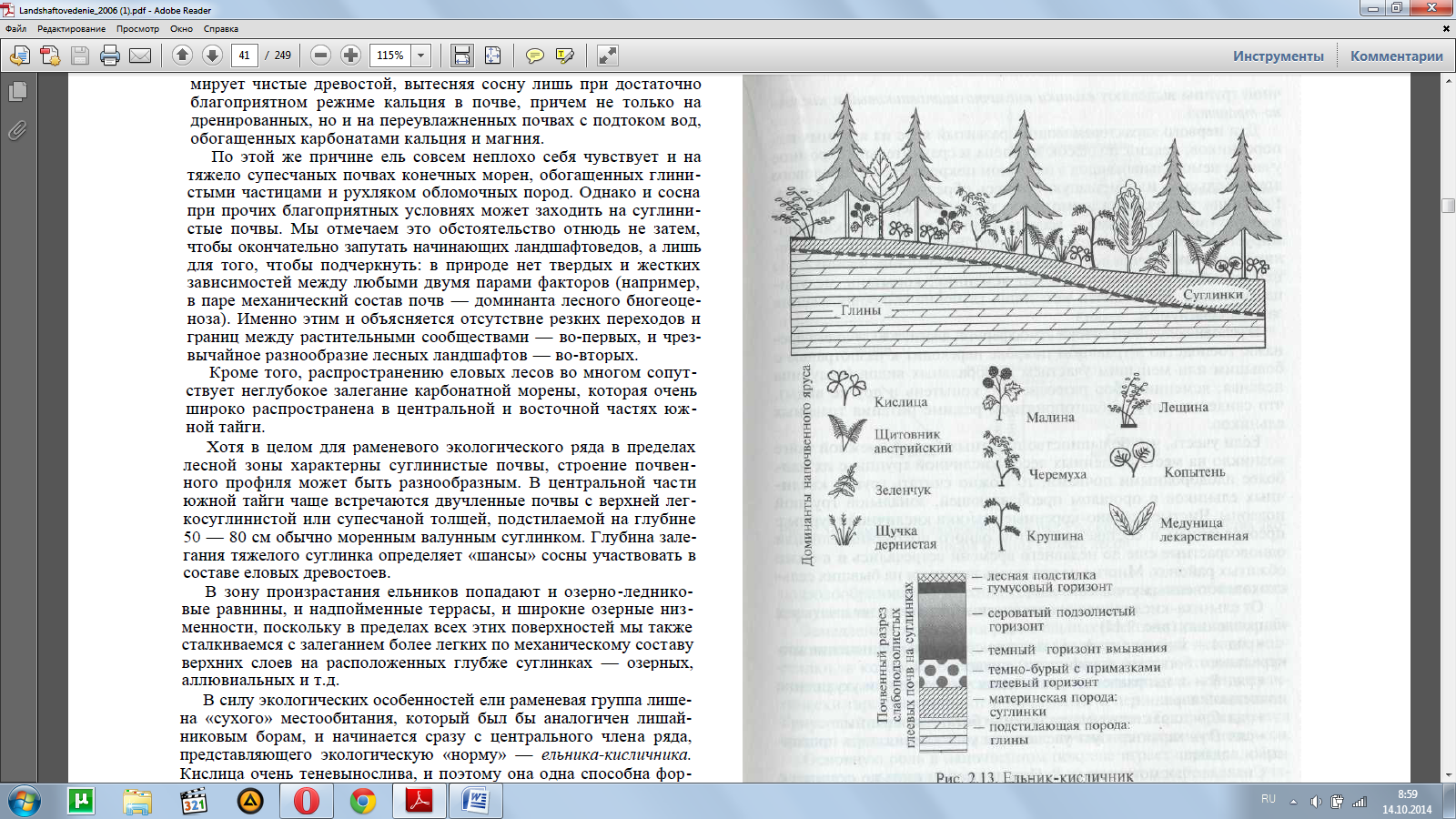
На глуби-не около 60 — 70 см залегает водоупорный тяжелый моренный суг-линок, над которым во влажные годы длительно задерживаетсяверховодка и развиваются анаэробные процессы, вызывающие оглеение. Следы бескислородного режима хорошо заметны: не-большие белесые пятна, марганцево-железистые мелкие конкре-ции. В связи с этим скелетная корневая система ели поверхност-ная и деревья в таких урочищах подвержены ветровалу.
В районах конечных морен и других возвышенных территорий преобладают леса кисличной группы, поэтому в пределах кисли-чной группы выделяют ельники кислично-щитовниковый и кислич-но-травяной.
Для первого характерен мощно развитый ярус из крупных па-поротников, редкий подлесок из клена и сравнительно скромное участие неморальных видов в наземном покрове. В составе елового яруса большую или меньшую примесь образуют осина и береза. Единично встречаются жимолость, крушина, черемуха, лещина; в «окнах» между кронами — малина. Щитовник австрийский фор-мирует мощный почти сплошной ярус. Травяной покров — кис-лица, майник, щучка дернистая, ожика волосистая, черника. Из неморальных видов присутствуют ясменник, копытень, медуница, зеленчук. Мхов почти нет, лишь глубокие микропонижения заняты сфагнумом.
В условиях лучшего оттока атмосферной влаги, т. е. лучшего дре-нажа, господство в травяном покрове переходит к разнотравью с большим или меньшим участием неморальных видов (медуница неясная, ясменник, бор развесистый, копытень и другие виды), что свидетельствует о благоприятном режиме питания травяных ельников.
Если учесть, что большинство пахотных угодий в южной тайге возникло на месте сведенных лесов кисличной группы с их наи-более плодородными почвами, то можно считать группу кисли-чных ельников в прошлом преобладающей, зональной группой подзоны. Чистые условно-коренные ельники кисличной группы с преобладанием в составе древостоя одного поколения ели или одновозрастные еще до недавнего времени встречались и в более обжитых районах. Многие из них вновь возникли на бывших сель-скохозяйственных угодьях.
От ельника-кисличника экологические ряды отходят в четырех направлениях (рис. 2.14):
• ряд А — характеризует увеличение сухости и уменьшение ми нерального богатства (трофности) почвы;
• ряд В — в направлении увеличения увлажнения и ухудшение почвенной аэрации;
• ряд С — характеризует увеличение богатства почв;
• ряд D — характеризует увеличение увлажнения почв проточ ными водами.

Сначала просмотрим ряд В. При движении вниз по склонам с увеличением увлажнения и одновременным ухудшения условий «дыхания» почв формируются урочища ельников-черничников (рис. 2.15).
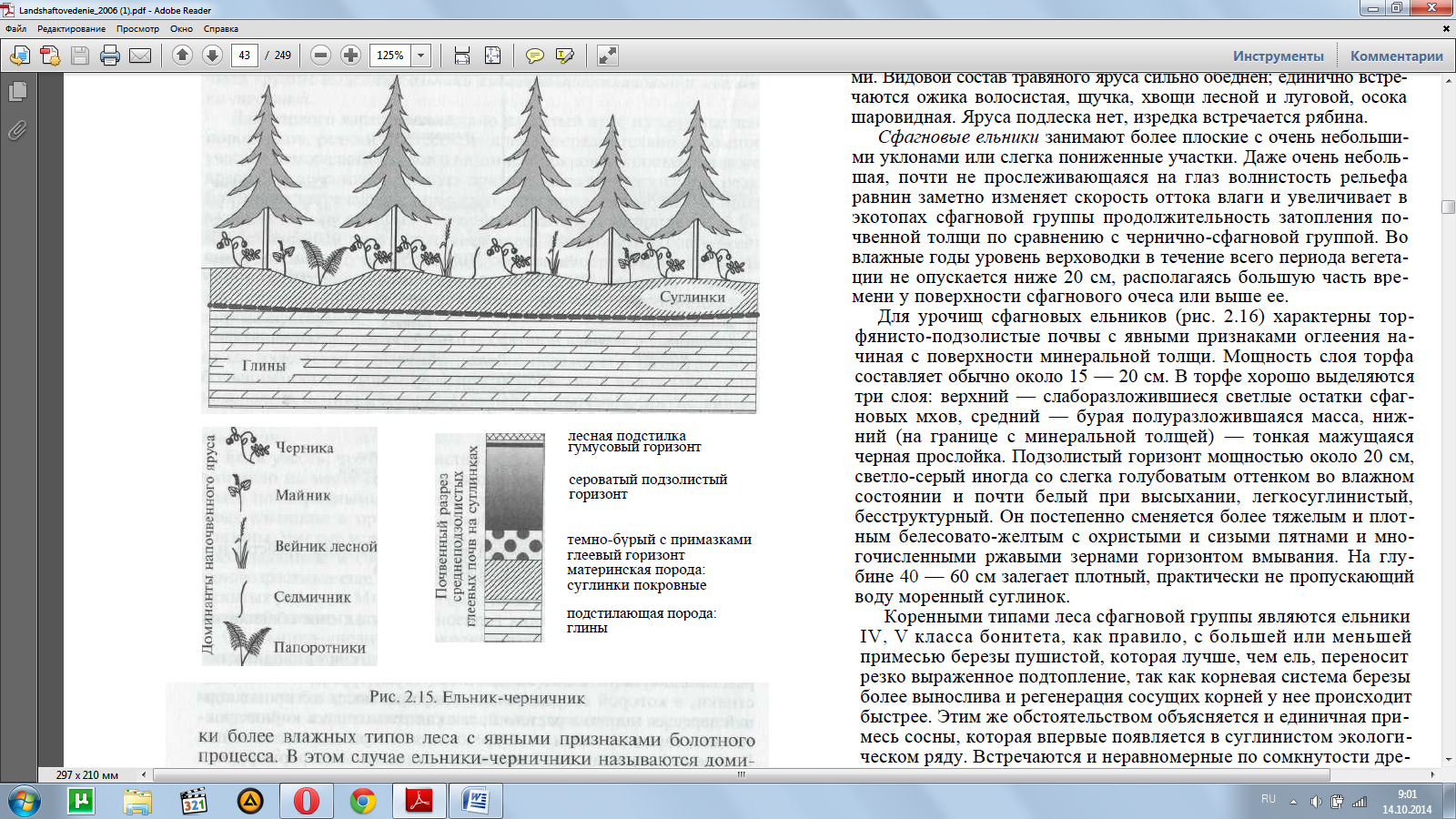
На более пологих склонах моренных равнин замедлен-ный отток влаги обусловливает подъем верховодки и периодичес-кое (весной, а во влажные годы и летом) затопление почвы до ее поверхности. Эта важнейшая особенность предопределяет направ-ленность почвообразования: при разложении растительных остат-ков возникают более кислые подвижные продукты, усиливаетсяподзолообразование верхних горизонтов почвы, они обедняются доступными элементами питания.
Замедление разложения растительных остатков приводит к об-разованию сравнительно мощной (3 — 6 см) грубой лесной под-стилки, в которой сосредоточена основная масса активных кор-ней деревьев и других растений, так как только здесь корни прак-тически гарантированы от повреждения в периоды подтопления. Гумусовый горизонт практически не выражен. Так формируются средне- и реже сильноподзолистые почвы ельников-черничников.
Основную роль в напочвенном покрове играет черника, обра-зующая в ельниках обычно сплошной покров с небольшим уча-стием вейника лесного, майника, седмичника, мелких папорот-ников и других видов таежного мелкотравья.
Волнистость и беспорядочная бугристость рельефа моренных равнин определяет мозаичность в распределении лесных урочищ, чутко реагирующих на изменение комплекса почвенных факто-ров, прежде всего режима аэрации и питания. Поэтому среди пре-обладающих по площади ельников-кисличников и ельников-чер-ничников вкраплены по плоским поверхностям и низинам участ-
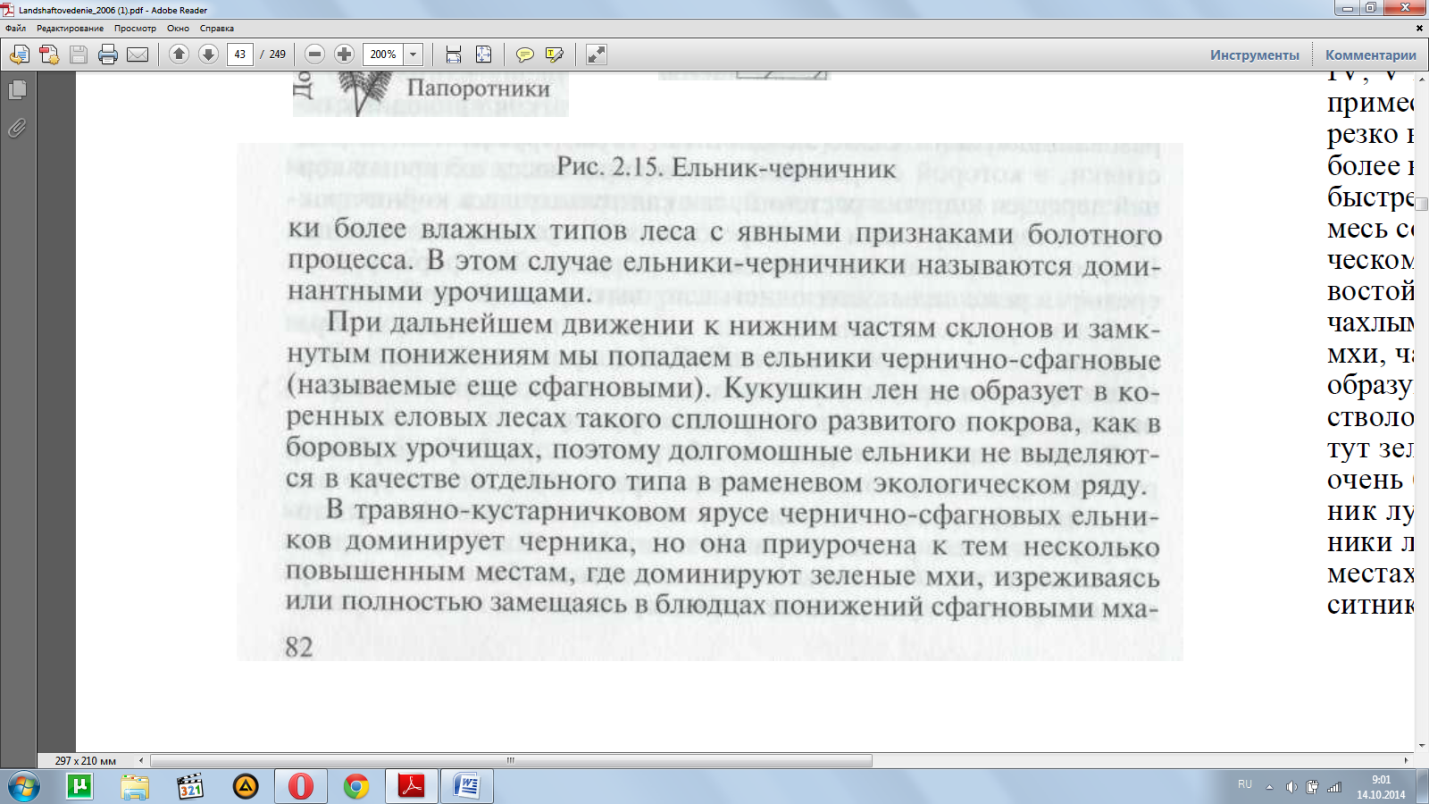
ми. Видовой состав травяного яруса сильно обеднен; единично встре-чаются ожика волосистая, щучка, хвощи лесной и луговой, осока шаровидная. Яруса подлеска нет, изредка встречается рябина.
Сфагновые ельники занимают более плоские с очень небольши-ми уклонами или слегка пониженные участки. Даже очень неболь-шая, почти не прослеживающаяся на глаз волнистость рельефа равнин заметно изменяет скорость оттока влаги и увеличивает в экотопах сфагновой группы продолжительность затопления по-чвенной толщи по сравнению с чернично-сфагновой группой. Во влажные годы уровень верховодки в течение всего периода вегета-ции не опускается ниже 20 см, располагаясь большую часть вре-мени у поверхности сфагнового очеса или выше ее.
Для урочищ сфагновых ельников (рис. 2.16) характерны тор-фянисто-подзолистые почвы с явными признаками оглеения на-чиная с поверхности минеральной толщи. Мощность слоя торфа составляет обычно около 15 — 20 см. В торфе хорошо выделяются три слоя: верхний — слаборазложившиеся светлые остатки сфаг-новых мхов, средний — бурая полуразложившаяся масса, ниж-ний (на границе с минеральной толщей) — тонкая мажущаяся черная прослойка. Подзолистый горизонт мощностью около 20 см, светло-серый иногда со слегка голубоватым оттенком во влажном состоянии и почти белый при высыхании, легкосуглинистый, бесструктурный. Он постепенно сменяется более тяжелым и плот-ным белесовато-желтым с охристыми и сизыми пятнами и мно-гочисленными ржавыми зернами горизонтом вмывания. На глу-бине 40 — 60 см залегает плотный, практически не пропускающий воду моренный суглинок.
Коренными типами леса сфагновой группы являются ельники IV, V класса бонитета, как правило, с большей или меньшей примесью березы пушистой, которая лучше, чем ель, переносит резко выраженное подтопление, так как корневая система березы более вынослива и регенерация сосущих корней у нее происходит быстрее. Этим же обстоятельством объясняется и единичная при-месь сосны, которая впервые появляется в суглинистом экологи-ческом ряду. Встречаются и неравномерные по сомкнутости дре-востой с большими лишенными деревьев прогалинами и совсем чахлыми елками. В напочвенном покрове господствует сфагновые мхи, часто с большим участием кукушкина льна; совместно они образуют живописный желто-зеленый мозаичный покров. На при-стволовых возвышениях, гниющем валеже, крупных кочках рас-тут зеленые мхи. Сюда же приурочено и большинство растений очень бедного по видовому составу травяного покрова: марьян-ник луговой, щучка, брусника, золотая розга, седмичник, вей-ники лесной и Лангсдорфа, иван-чай. На ровных и пониженных местах по покрову мхов встречаются лишь единичные растения ситника развесистого, хвоща лесного.

Рис. 2.16. Ельник сфагновый
В очень изреженном подлеске встречаются рябина; ивы уша-стая и пепельная по наиболее переувлажненным участкам (где березовый древостой изрежен) образуют довольно густой подле-сок высотой 1,0— 1,5 м. Елового подроста мало, и распределен он очень неравномерно, располагаясь, главным образом, на микро-повышениях.
Ельники сфагновые почти не сохранились в южной тайге, встре-чаются лишь в более глухих районах.
Урочища сфагновой группы на суглинистых торфянисто-гле-евых почвах можно рассматривать как начальный этап болотооб-разования, который в дальнейшем, по мере нарастания торфа, неизбежно приведет к еще большему переувлажнению, уменьше-нию трофности почвогрунтов и к смене ельников заболоченными сосняками, а затем и сфагновыми болотами.
Итак, мы рассмотрели типы раменевых урочищ суглинистых отложений равнинной тайги, для которых характерно увлажне-ние за счет выпадающих непосредственно на данную площадь ат-мосферных осадков, а отток влаги происходит, главным образом, по начальным элементам гидрографической сети — ложбинам и лощинам, временным водостокам, заполняющимся лишь талой водой или после обильных дождей.
Но существует весьма разнообразные экотопы, которым свой-ственно присутствие влаги, перемещающейся с соседних участ-ков внутри или по поверхности почвы и обогащенной питатель-ными элементами. Рельеф тяжелосуглинистых отложений основ-ной морены осложнен малозаметными узкими или довольно ши-рокими корытоообразными ложбинами, по которым идет сток вод весной и при обильных летних дождях. Эти ложбины в низо-вых частях сливаются друг с другом, и при пологом рельефе могут формироваться более или менее широкие полосы, а в низинах и значительные участки биогеоценозов проточного увлажнения с зарослями таволги, с болотным высокотравьем или покровом из сфагновых мхов.
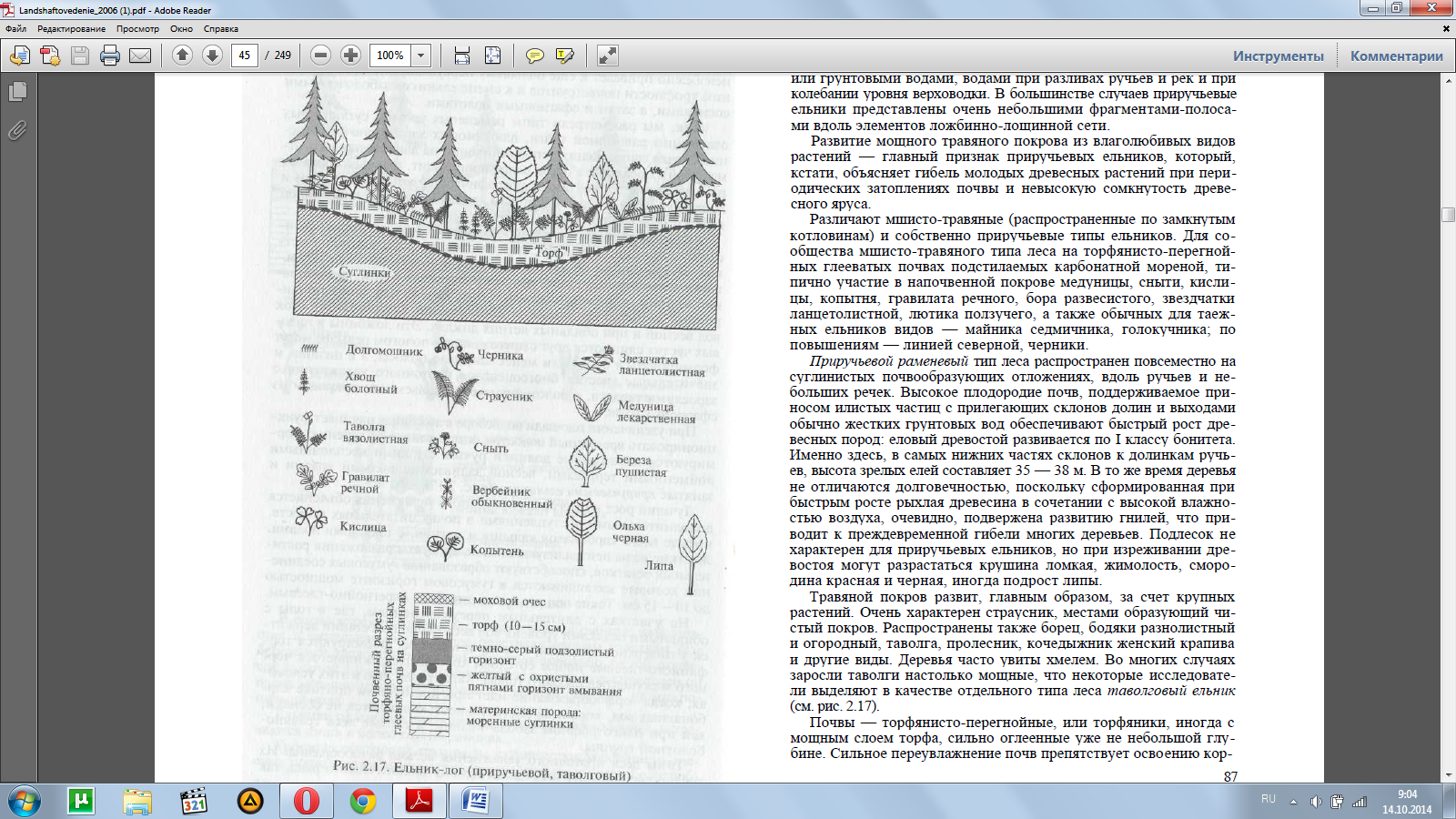
При увеличении площади водосбора в ложбинах начинает функ-ционировать временный водосток, а затем ниже по течению фор-мируются выраженные долинки ручьев с узкими несплошными пойменными террасами, весной заливаемые полыми водами и занятые приручьевыми ельниками (рис. 2.17).
Лучший рост в типах леса проточного увлажнения объясняется дополнительными поступлениями в почву питательных веществ, прежде всего карбонатов кальция и магния, с текущими водами. Жесткие воды нейтрализуют кислые продукты разложения расти-тельных остатков, способствуют образованию гумусовых соедине-ний которые накапливаются в гумусовом горизонте мощностью до 10—15 см. Такие почвы можно отнести к перегнойно-глеевым. На участках с длительным переувлажнением, где в годы с обильными осадками уровень вод весь период вегетации держится у поверхности почвы, а часто и выше ее, формируются тор-фянисто-глеевые почвы со слоем хорошо разложившегося чер-ного мажущегося рыхлого торфа. Характерно, что в этих услови-ях, когда торфообразование идет при постоянном притоке кар-бонатных вод, коренной растительностью являются не сосняки, как при олиготрофном заболачивании, а еловые леса травяно-болотной группы.
Типы леса проточного увлажнения весьма разнообразны. Их трудно расположить в пределах одного экологического ряда, так
как основная особенность — увлажнение обогащенными минера-лами водами — достигается разными путями: с поверхностными или грунтовыми водами, водами при разливах ручьев и рек и при колебании уровня верховодки. В большинстве случаев приручьевые ельники представлены очень небольшими фрагментами-полоса-ми вдоль элементов ложбинно-лощинной сети.
Развитие мощного травяного покрова из влаголюбивых видов растений — главный признак приручьевых ельников, который, кстати, объясняет гибель молодых древесных растений при пери-одических затоплениях почвы и невысокую сомкнутость древе-сного яруса.
Различают мшисто-травяные (распространенные по замкнутым котловинам) и собственно приручьевые типы ельников. Для со-общества мшисто-травяного типа леса на торфянисто-перегной-ных глееватых почвах подстилаемых карбонатной мореной, ти-пично участие в напочвенной покрове медуницы, сныти, кисли-цы, копытня, гравилата речного, бора развесистого, звездчатки ланцетолистной, лютика ползучего, а также обычных для таеж-ных ельников видов — майника седмичника, голокучника; по повышениям — линией северной, черники.
Приручьевой раменевый тип леса распространен повсеместно на суглинистых почвообразующих отложениях, вдоль ручьев и не-больших речек. Высокое плодородие почв, поддерживаемое при-носом илистых частиц с прилегающих склонов долин и выходами обычно жестких грунтовых вод обеспечивают быстрый рост дре-весных пород: еловый древостой развивается по I классу бонитета. Именно здесь, в самых нижних частях склонов к долинкам ручь-ев, высота зрелых елей составляет 35 — 38 м. В то же время деревья не отличаются долговечностью, поскольку сформированная при быстрым росте рыхлая древесина в сочетании с высокой влажно-стью воздуха, очевидно, подвержена развитию гнилей, что при-водит к преждевременной гибели многих деревьев. Подлесок не характерен для приручьевых ельников, но при изреживании дре-востоя могут разрастаться крушина ломкая, жимолость, сморо-дина красная и черная, иногда подрост липы.
Травяной покров развит, главным образом, за счет крупных растений. Очень характерен страусник, местами образующий чи-стый покров. Распространены также борец, бодяки разнолистный и огородный, таволга, пролесник, кочедыжник женский крапива и другие виды. Деревья часто увиты хмелем. Во многих случаях заросли таволги настолько мощные, что некоторые исследовате-ли выделяют в качестве отдельного типа леса таволговый ельник (см. рис. 2.17).
Почвы — торфянисто-перегнойные, или торфяники, иногда с мощным слоем торфа, сильно оглеенные уже не небольшой глу-бине. Сильное переувлажнение почв препятствует освоению кор-
нями деревьев не только более глубоких слоев почвы, но в пони-женных местах и самого верхнего слоя, в результате чего создают-ся «мертвые» для корней деревьев зоны, занимающие большую или меньшую часть всей поверхности в зависимости от общего режима переувлажнения данного участка.