

875
.pdf4.Для отбора маточников с высоким выходом семян достаточно определить их содержание всего в 3–4 шишках, которое высоко коррелирует с их выходом в 20–30 шишках.
5.При выделении особей с близким к нулю выходом семян их следует исключать при наборе клонов для обычных ЛСП, где опыление будет перекрестным. Однако их следует обязательно сохранить в архиве клонов как особо ценный материал для сорта-самоопылителя.
Контрольные вопросы:
1. Влияние размера шишек и семян сосны на рост семей. Частота лучших семей по градациям этих признаков в среднем и колебания по отдельным опытам. Динамика корреляций в возрасте семей от 3 до 18 лет. Движущий (рис. 6.2) и стабилизирующий (рис. 6.3) отбор
2. Влияние темной, пестрой, коричневой и светлой окраски семени на рост потомства.
Частота лучших семей по градациям окраски: а) в среднем по многим опытам; б) колебания по отдельным популяциям. Влияние окраски на время прорастания семян.
3. Вклад клонов в генный пул семян по В.В. Тараканову. Рекомендации по селекции на урожайность: показать разную повторяемость выхода семян и числа шишек у материнских деревьев, и почему возможен отбор урожайных деревьев по минимальной выборке. Зачем следует сохранять в архиве клонов деревья с близким к нулю выходом семян.
190
Глава 7. КОРРЕЛЯТИВНАЯ СЕЛЕКЦИЯ ЕЛИ
7.1. Влияние размера шишек и массы семени на рост семей
Длина шишек ели является признаком, определяющим ее видовую принадлежность. На европейском Северо-Западе и Урале расположена зона естественной интрогрессивной гибридизации двух видов ели: европейской и сибирской. Исследования (Попов, 2005) показали, что здесь образуется множество переходных форм с преобладанием признаков ели сибирской ближе к Уралу и северу и признаков ели европейской к западу и югу. Обширность зоны гибридизации позволила даже систематикам отнести эти формы к новому виду и назвать ее ель гибридная или финская (Picea fennica (Regel) Kom.)
(Правдин, 1975).
При анализе шишек и семян обращалось внимание на разную длину шишек в 3 совокупностях: плюс-деревьев естественных популяций, плюсдеревьев из лесных культур и обычных деревьев из лесных культур. При этом исключили популяцию «Нижняя Курья», так как культуры в ней были созданы гораздо позднее (в 1939 г.), чем на 4-х других участках, которые создавались в 1901–1916 гг. под руководством Ф.А. Теплоухова. Но не только поэтому.
Вариация длины шишек у плюс-деревьев колебалась от 8 до 15.5%, составляя в среднем в естественных насаждениях 11.6% и в культурах 10.3%. Минимум и максимум длины шишек в естественных насаждениях составили 60 и 111 мм, а в культурах 65 и 106 мм, т.е. были близки. Но в популяции «Нижняя Курья» эти колебания составили 64–85 мм при средней длине в выборке плюсдеревьев 76.1 мм и в выборке обычных деревьев 73.3 мм, что оказалось значительно меньше, чем в культурах Ф.А. Теплоухова (происхождения «Сепыч 1», «Сепыч 2», «Очер» и «Верещагино» (табл. 7.1).
Средняя длина шишек у плюс-деревьев в 7 естественных популяциях оказалась 81.2 мм, а в 4 ценопопуляциях культур Ф.А. Теплоухова 87.9 мм или на 8.2% больше. В целом обе эти группы можно отнести к одному виду (подвиду) ели – ели гибридной (финской), у которой средняя длина шишек определена 85±6.5 мм (Правдин, 1975). Средняя масса семени и число семян в шишке также оказались больше в культурах, на 8.5% и 23.7% соответственно. Достоверности различий оказались выше стандартных при tф= 7.1 для длины шишек, tф= 4.1 для массы семени и tф= 4.7 для числа семян в шишке.
Вероятно, при закладке культур под руководством Ф.А. Теплоухова были использованы семена, сознательно заготовленные от деревьев с более
191
крупными шишками. В свое время Федор Александрович Теплоухов интересовался этим вопросом и опубликовал самую первую статью о морфометрии шишек ели в европейской части России (Теплоухов, 1876).
Табл.7.1 – Морфометрия шишек и семян ели финской в Пермском крае
|
Число |
Длина шишек |
Масса семени |
Число семян в |
1 |
|||||
|
шишке |
|
|
|||||||
Происхождение |
дере- |
|
|
|
|
|
|
|
||
|
Вари- |
|
|
Вари- |
|
|
Вари- |
|||
(ценопопуляция) |
вьев, |
Среднее, |
Среднее |
|
Среднее, |
|
||||
ация, |
|
ация, |
|
|
ация, |
|||||
|
шт |
мм |
0,1 мг |
|
шт. |
|
|
|||
|
% |
|
% |
|
|
% |
||||
|
|
|
|
|
|
|
|
|||
|
Естественные насаждения, |
плюсовые деревья |
|
|
|
|
||||
Очер |
13 |
87,0±1,93 |
8,0 |
55,8±2,25 |
|
14,6 |
95,7±11,3 |
|
|
42,6 |
Нытва |
29 |
84,4±2,15 |
13,7 |
51,4±1,67 |
|
17,5 |
80,7±7,5 |
|
|
49,9 |
Гайва |
74 |
82,2±1,16 |
12,2 |
53,3±0,93 |
|
15,1 |
110,0±4,0 |
|
|
31,2 |
Ильинск |
32 |
82,3±1,93 |
13,3 |
53,8±1,85 |
|
19,5 |
96,6±6,5 |
|
|
37,9 |
Пермь |
108 |
78,2±0,92 |
12,2 |
50,0±0,73 |
|
15,2 |
92,5±3,8 |
|
|
42,6 |
Кунгур |
27 |
77,3±1,69 |
11,3 |
48,7±1,78 |
|
19,0 |
65,2±7,1 |
|
|
56,3 |
Чусовой |
18 |
77,3±1,91 |
10,5 |
51,4±2,21 |
|
18,3 |
80,0±6,3 |
|
|
33,1 |
В среднем |
|
81,24 |
11,6 |
52,07 |
|
17,0 |
88,7 |
|
|
42,0 |
Вся совокупность |
301 |
80,4±0,59 |
12,6 |
51,6±0,50 |
|
16,7 |
93,1±2,25 |
|
|
41,9 |
|
Лесные культуры, плюсовые деревья |
|
|
|
|
|
||||
Сепыч-I |
85 |
87,6±0,94 |
9,9 |
54,9±0,86 |
|
14,3 |
112,6±4,5 |
|
|
36,9 |
Сепыч-II |
9 |
92,3±2,85 |
9,2 |
61,8±3,01 |
|
14,6 |
100,0±11,3 |
|
|
33,9 |
Очер |
16 |
84,3±2,54 |
12,0 |
54,3±1,59 |
|
11,7 |
109,3±8,3 |
|
|
30,4 |
Верещагино |
19 |
87,5±2,01 |
10,0 |
55,0±2,24 |
|
17,7 |
116,8±6,4 |
|
|
23,7 |
Н. Курья |
23 |
76,1±0,89 |
5,6 |
54,6±1,70 |
|
15,0 |
118,9±8,9 |
|
|
35,7 |
В среднем без Н.Курьи |
87,92 |
10,3 |
56,48 |
|
14,6 |
109,7 |
|
|
31,2 |
|
Вся совокупность без |
|
|
|
|
|
|
|
|
|
|
Н.Курьи |
129 |
87,46±0,79 |
10,2 |
55,24±0,72 |
|
14,9 |
112,0±3,37 |
|
|
34,2 |
Лесные культуры, обычные деревья (нормальные и минусовые) |
|
|
||||||||
Сепыч-I |
19 |
81,7±1,50 |
8,0 |
53,8±1,53 |
|
12,4 |
119,5±9,5 |
|
|
34,6 |
Сепыч-II |
15 |
83,0±2,49 |
11,6 |
56,4±2,89 |
|
19,9 |
104,6±7,4 |
|
|
27,2 |
Очер |
13 |
83,5±3,58 |
15,5 |
49,9±3,17 |
|
22,9 |
126,2±8,3 |
|
|
23,9 |
Верещагино |
11 |
79,6±2,09 |
8,7 |
51,5±1,97 |
|
12,6 |
130,0±13,0 |
|
|
33,2 |
Н. Курья |
14 |
73,3±2,20 |
11,2 |
51,7±1,95 |
|
14,1 |
130,9±12,5 |
|
|
35,6 |
В среднем без Н.Курьи |
81,9 |
11,0 |
52,9 |
|
17,0 |
120,1 |
|
|
29,7 |
|
Вся совокупность без |
|
|
|
|
|
|
|
|
|
|
Н.Курьи |
58 |
82,23±1,19 |
11,0 |
53,05±1,21 |
|
17,4 |
116,8±42,0 |
|
|
27,3 |
|
|
|
|
|
|
|
|
|
|
|
Распределение плюс-деревьев естественных популяций имеет два модальных класса длины шишек в пределах 71–80 мм. Распределение шишек
192
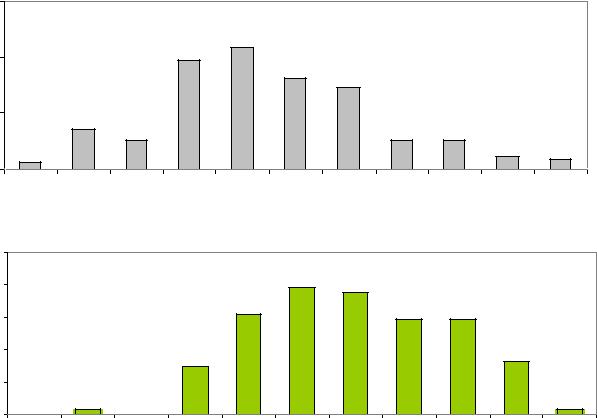
плюс-деревьев из культур заметно отличается общим сдвигом вправо и почти полным отсутствием шишек длиной менее 70 мм, а также двумя модальными классами в пределах 81–90 мм (рис. 7.1).
30
Частота, %
20 |
|
|
|
|
|
|
10 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
56-60 |
61-65 |
66-70 71-75 |
76-80 81-85 86-90 |
91-95 96-100 |
101-105 |
106-110 |
|
|
Длина шишек ели в естественных популяциях, мм |
|
|
||
25
Частота, %
20 |
|
|
|
|
|
|
15 |
|
|
|
|
|
|
10 |
|
|
|
|
|
|
5 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
56-60 |
61-65 |
66-70 71-75 |
76-80 81-85 86-90 |
91-95 96-100 |
101-105 |
106-110 |
|
|
Длина шишек в культурах Теплоухова, мм |
|
|
||
Рис. 7.1 – Длина шишек ели в естественных популяциях и в культурах Ф.А. Теплоухова
Поэтому в культурах деревья по длине шишек можно отнести к форме с преобладанием признаков ели европейской, а в естественных популяциях – к форме с преобладанием признаков ели сибирской.
В культурах отмечены и другие особенности, которые можно отнести к критериям, по которым одна популяция отличается от другой. В частности, в ценопопуляциях «Сепыч 1» и «Очер» встретились 4 плюсовых дерева с гладкой корой (3.5% деревьев). Это своего рода «фен», который свидетельствует о том, что мы имеем дело с двумя группами популяций. К первой относятся культуры, где фен гладкой коры есть, а ко второй – естественные насаждения, где такого фена нет. Наличие этого фена может говорить и о другом – он проявился только в связи с заготовкой семян в какой-то отдельно расположенной и, скорее
193
всего, более юго-западной популяции (но в пределах бывшего Очерского окружного лесничества в имении графов Строгановых). Во всяком случае, есть все основания для выделения «культур Теплоухова» по этим признакам (длине шишек и форме коры) в отдельные ценопопуляции, которые были образованы искусственно в результате первого приема массового отбора деревьев с более длинными шишками.
Деревья с такими более длинными шишкам условно можно считать носителями «европейских», а с короткими – «сибирских» генов ели. Тогда в случае группового расположения потомств (вицинизма) возможна эволюция с образованием ценопопуляций более «сибирских» и более «европейских», чем исходная популяция. Кроме того, пул генов в таких потомствах будет формировать какие-то новые сочетания, обусловленные большей фертильностью и урожаем пыльцы отдельных деревьев. Именно они сформируют специфику генофонда и будут причиной появления разных тенденций отбора в современной эволюции этого гибридного вида ели.
По классификации Л.Ф. Правдина (Правдин, 1975), у ели европейской средняя длина шишек определена 100±10 мм, ели сибирской 62±10 и ели гибридной или финской 85±6.5 мм. Если придерживаться этих параметров, хотя имеются и иные сведения (Попов, 2005), то частота встречаемости плюсовых деревьев с шишками европейского типа (от 90 мм и более) в естественных насаждениях и культурах составила, соответственно, 14.5 и 38.2% (в культурах в 2.6 раза больше). С другой стороны, встречаемость деревьев с шишками сибирского типа (до 72 мм) в природных популяциях 18.9%, а в культурах 5.7%. То есть «сибирских» плюс-деревьев оказалось больше в 3.3 раза в естественных популяциях.
Возникает закономерный вопрос, а как же растут «европейские» и «сибирские» деревья в этих совокупностях? В естественных насаждениях мы имеем выборки только из плюсовых деревьев, но в культурах есть выборки и из обычных деревьев. Объединение их в культурах Ф.А. Теплоухова и дает ответ на поставленный вопрос.
В этой объединенной выборке из 129 плюсовых и 58 обычных деревьев между высотой деревьев и длиной шишек была обнаружена слабая корреляция
194
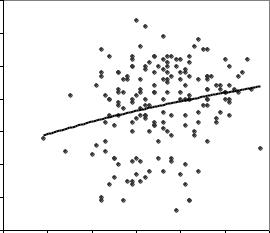
(r = 0.21±0.07). То есть более «европейские» деревья ели в ней растут лучше и
оказываются выше (рис. 7.2.). |
|
|
|||||
|
150 |
|
|
|
|
|
|
|
140 |
|
|
|
|
|
|
% |
130 |
|
|
|
|
|
|
, |
|
|
|
|
|
|
|
деревьев |
120 |
|
|
|
|
|
|
110 |
|
|
|
|
|
|
|
Высота |
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
90 |
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
50 |
60 |
70 |
80 |
90 |
100 |
110 |
|
|
Длина шишек в культурах Теплоухова, мм |
|
||||
Рис. 7.2 – Связь между длиной шишек ели и высотой материнских деревьев в культурах Ф.А. Теплоухова.
Вприродных популяциях такой связи нет (r=0.004). Тогда можно полагать, что гены ели европейской, которых оказалось больше в культурах, способствуют тому, что их обладатели развиваются в более крупные деревья.
Если принять во внимание некоторую пространственную изоляцию ценопопуляций культур и формирование пула семян при опылении их же пыльцой, то тогда после отбора «крупношишечных матерей», проведенного в начале 20 века Ф.А. Теплоуховым, мы наблюдаем сейчас в потомствах F1 и F2 следующие результаты.
Впотомстве F1 (культуры Ф.А. Теплоухова) получено увеличение: длины шишек на 8.2%, массы семени на 8.5% и высоты культур на 1 класс бонитета. Увеличение высоты культур произошло в результате действия комплекса причин, однако и хорошее качество семян было одной из них.
Потомство F2 (потомство плюс-деревьев из культур) показало высоту 106.6% при высоте потомства естественных насаждений 102.4% (преимущество F2 +4.2%), причем потомство нормальных и минусовых деревьев из культур превышало контроль на 1.0–5.5% (см. табл. 7.1).
Далее анализ полей корреляции показал еще много интересного. В предыдущих разделах мы эффективно использовали метод анализа долей. Этот метод весьма чувствителен и часто дает ответы на селекционные вопросы, когда корреляции близки к нулевым значениям.
195
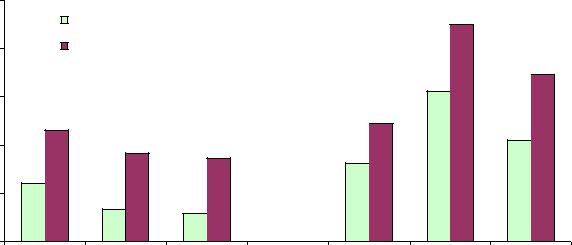
Оказалось, что в культурах Ф.А. Теплоухова, т.е. в более «европейских» ценопопуляциях, у матерей со средней массой семени лучших семей с высотой от 115% (левые светлые столбики) оказалось в 1.9 и 1.5 раза больше, чем у матерей с легкими и с тяжелыми семенами. Следовательно, в отношении массы семени в культурах действует стабилизирующий отбор (рис. 7.3).
Доля лучших семей, %
50 |
|
|
|
45,1 |
|
|
лучшие от 115% |
|
|
|
|
|
|
|
|
|
|
40 |
лучшие от 110% |
|
|
|
|
|
|
|
|
|
34,6 |
|
|
|
|
31,4 |
|
30 |
|
|
|
|
|
23,1 |
|
|
24,5 |
|
|
|
|
|
|
21,2 |
|
|
|
|
|
|
|
20 |
18,3 |
17,3 |
16,3 |
|
|
|
|
|
|||
|
|
|
|
|
|
12,1 |
|
|
|
|
|
10 |
6,7 |
5,8 |
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
легкие |
средние |
тяжелые |
легкие |
средние |
тяжелые |
Естественные насаждения |
Культуры Теплоухова |
|
|||
Рис. 7.3 – Доля лучших семей |
в 21-летнем потомстве |
ели из |
естественных насаждений (столбики слева, |
движущий отбор) и из |
культур |
Ф.А. Теплоухова (столбики справа, стабилизирующий отбор) по градациям массы семени у материнских деревьев (легкие, средние и тяжелые семена).
Однако в естественных насаждениях, т.е более «сибирских», преимущество изменилось в пользу легких семян, где лучших семей оказалось 12.1%, а у деревьев со средними и тяжелыми – 6.7 и 5.8%, т.е. больше в 1.8–2.1 раза. Таким образом, здесь мы наблюдаем уже другой вид естественного отбора – отбор движущий. Различия сохраняются также и при отборе лучших семей от 110% (правые темные столбики).
Эти различия в тенденциях отбора и возможной эволюции у ели финской, насколько можно судить по сведениям из литературы, обнаружены впервые.
На следующих двух полях корреляции, приведенных ниже по данным для плюс-деревьев из естественных насаждений, треугольными точками выделены 6 быстрорастущих семей (рис. 7.4).
Этот кластер образуют семьи 6 плюс-деревьев из самых разных популяций: Нытвы, Ильинска, Кунгура и Перми. У этих 6 деревьев легкие
196
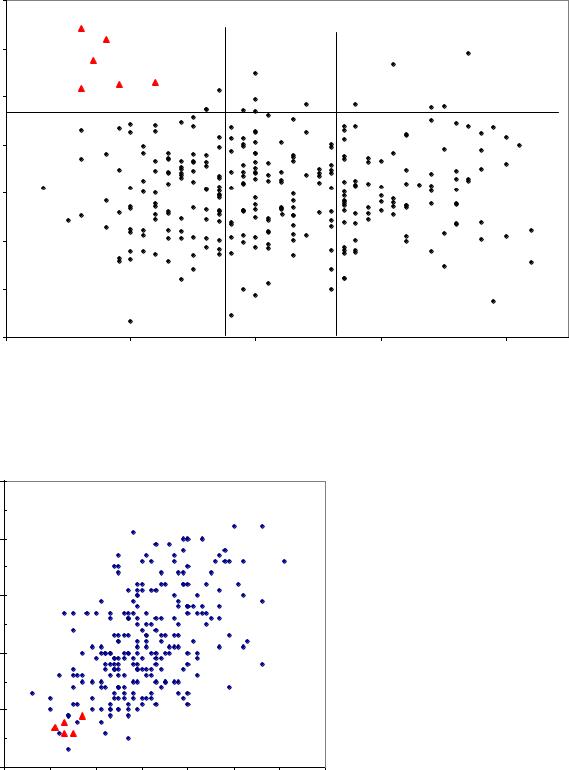
семена сочетаются с короткими шишками, т.е. по классификации Л.Ф. Правдина (1975), это типичные «сибирские» ели, для которых средняя длина шишек определена 62±10 (рис. 7.5).
Высота потомства в 21 год, %
140
130
120
110
100
90
80
70
30 |
40 |
50 |
60 |
70 |
Масса семени плюс-деревьев из естественных популяций, 0001 г
Рис. 7.4 – Масса семени и высота 21-летнего потомства у плюс-деревьев ели из естественных популяций. Пояснения в тексте
Масса семени, 0,0001 г
80
70
60
50
40
30
50 |
60 |
70 |
80 |
90 |
100 |
110 |
120 |
Длина шишиек, мм
Рис. 7.5 – Связь длины шишек и массы семени у плюс-деревьев ели из естественных популяций. Пояснения в тексте.
197
Но даже если обнаруженный кластер исключить, то доли лучших семей в трех градациях массы семени, обозначенных вертикальными линиями на поле корреляции, просто уравняются. В таком случае для трех видов естественного отбора – стабилизирующего, движущего и дизруптивного здесь ситуация такова, что если для ели финской из естественных популяций вдруг потребуется эволюция в сторону быстрорастущих семей (человек эту эволюцию делает искусственно, называя селекцией), то можно ожидать у ели появление все большего числа потомков с мелкими шишками и семенами.
С другой стороны, остаются некоторые сомнения в части возможного влияния на размер шишек более молодого возраста деревьев, который в культурах Ф.А. Теплоухова был 74–88 лет, а естественные популяции были старше: 90–143 года. Однако один из участков культур, не относящийся к культурам Теплоухова (Нижне-Курьинский) был самым молодым (51 год). Тем не менее, в нем длина шишек и масса семени были наименьшими, а высота семей оказалась самой низкой. Поэтому молодой возраст деревьев не является тем фактором, который увеличивает размеры шишек и семян.
При описании методики выделения плюс-деревьев уже шла речь о том, что на популяции было давление отбора с изъятием продуктивных растений и это отразилось на качестве плюсовых деревьев, отобранных вблизи г. Перми (происхождения Пермь и Гайва). Однако в популяциях Очер, Нытва и Ильинск такого давления не было, однако и в них есть деревья, входящие в кластер с малой массой семени. Поэтому можно дать следующее объяснение движущего отбора в пользу деревьев с самыми мелкими шишками и семенами.
В изученных естественных популяциях ели финской (гибридной) преобладают формы с признаками ели сибирской, поэтому можно полагать, что протекающие в них процессы образования семян идут иным образом, чем в ценопопуляциях культур, где был проведен первый отбор крупношишечных форм, причем это было сделано, возможно, в наиболее продуктивных ее популяциях с преобладанием форм ели европейской. Поэтому и тренды эволюции этих форм ели финской различны, что и показали результаты изучения их потомства.
198
Более «европейская» ель (в культурах Ф.А. Теплоухова) на селекцию крупношишечных форм реагировала стабилизацией потомства F2, с преимуществом в росте потомства от деревьев со средними семенами. Т.е. популяции вернулись в устойчивое состояние и продолжение отбора на большой размер шишек бесперспективен. Вообще, на такие же результаты массового отбора указывают многочисленные примеры среди других видов растений, когда эффективен только его начальный этап.
У более «сибирской» формы ели в природных популяциях такой стабилизации нет, и ее селекция почти равнозначна и по крупным, и по мелким шишкам и семенам. В ее потомствах неожиданно проявилась слабая тенденция к движущему отбору в отношении деревьев с малой массой семени и, как следствие, возможному появлению в следующих поколениях все большего числа потомков с мелкими семенами.
Часто лесные генетики опасаются сокращения генетического разнообразия в лесных культурах. Но селекция также «сократитель» биоразнообразия, так как изымает из генофонда только ту его часть, которая кажется человеку лучшей, а часть не может быть больше целого. Естественный отбор в одном поколении – это «шаги» эволюции. Их надо выявить и использовать в селекции – эволюции, направляемой человеком. Если бы природе потребовались быстрорастущие семьи ели, то ее эволюция в сходных с испытательными культурами условиях была бы направлена на выживание матерей с мелкими семенами, а эволюция потомства из культур – на матерей со средними семенами.
Именно такие разные родители от ели финской и требуются человеку, и необходимо использовать обнаруженное явление в ее уже искусственной эволюции, т.е. селекции.
Выводы.
1. Ель в культурах Ф.А. Теплоухова относится к форме с преобладанием признаков ели европейской, а в естественных популяциях – к форме с преобладанием признаков ели сибирской. Различия основаны на длине шишек у плюс-деревьев, которые в культурах оказалась на 8.2% длиннее; масса семени
199