228 |
C h a p t e r 2 3 : THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
Figure 23–7. Left lateral view of the brain and skull. At this stage, as during the previous 5 weeks, the brain dominates the morphological appearance of the head. The chondrocranium is mainly a basal support for certain parts: the surface of the spheno-ethmoidal cartilage for the anterior part of the olfactory bulb; the sphenoid for the hypothalamus and the hypophysis. The mesencephalon and the cerebellum, however, lack such a basal cartilaginous support. The pons is buttressed mediobasally by the basisphenoid and laterobasally by the cochlear portion of the otic capsule. The portions of the eye and brain covered by skeletal elements are shown by interrupted lines.
The protective covering of the superolateral surface of the cerebral hemispheres is beginning to be strengthened by the frontal bones. The lateral portions of the cerebellum and of the pons and medulla oblongata are encased by the parietal laminae and by the tectum posterius and pilae occipitales. The foramen magnum is very extensive, portions of the medulla oblongata being surrounded by mesenchyme only. The cartilages of the pharyngeal arches, including the first (Meckel’s), are well developed: those for the auditory ossicles, styloid process, and hyoid and laryngeal cartilages (Muller,¨ O’Rahilly, and Tucker, 1981). The neural arches of the vertebrae have not fused across the median plane. The normal spina bifida totalis characteristic of stage 23 has been studied in detail (O’Rahilly, Muller,¨ and Meyer, 1980, 1983, 1990a,b).
THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
229 |
r
Figure 23–8. Dorsal (superior) schematic views of the chondrocranium at stages 22 and 23, drawn to the same scale.
(A)The main components of the skull are listed on the left, the chief foramina and future canals on the right. At the junction of the basisphenoid, basioccipital, greater wing of the sphenoid, and petrous temporal, the carotid foramen is visible (in black). A superimposed outline (double line) of the chondrocranium at stage 23 aids in showing the considerable increase in size between A and B within 2 or 3 days. (B) The internal acoustic foramen (future meatus) is again evident in the petrous temporal, the jugular foramen is situated posteriorly, and the hypoglossal foramen lies close to the foramen magnum. The thick black, vertical line in both A and B is the notochord in transit from the dens to the cranial base. The main changes are the acquisition of the nasal capsule, the widening of the greater wings of the sphenoid (which support the cerebral hemispheres), and the formation of the parietal laminae (which include the foramen magnum). The occipital part of the brain sits in a skeletal bowl, from which, however, it is protected by fluid-filled mesenchyme (as in the area between the rhombencephalon and the basicranium in Fig. 23–6). The inventory of the “membrane” bones is almost complete in some embryos of stage 23.
230 |
C h a p t e r 2 3 : THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
Dural limiting layer
 Falx cerebri
Falx cerebri
Medial wall of  right cerebral
right cerebral
hemisphere
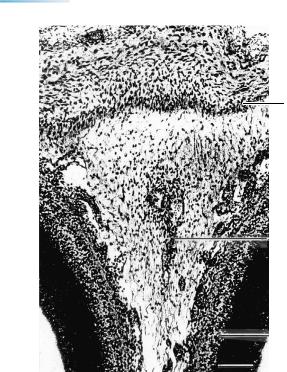
Figure 23–9. The dura mater, silver-impregnated. The falx cerebri is clearly formed from the skeletogenous layer, beginning at the level of the future crista galli. Most of the tissue between the cerebral hemispheres is still leptomeningeal, whereas the dural limiting layer is fibrous. The primordium of the superior sagittal sinus is visible at the edge of the developing falx. The beginnings of the transverse and occipital sinuses are also present and are intradural. See key on next page. Bar: 0.2 mm.
THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
231 |
|
Skeletogenous |
|
|
|
layer |
|
|
|
Dural limiting |
Subarachnoid |
|
|
space |
|
|
layer |
|
|
|
|
|
|
Lateral |
|
|
|
forebrain |
Lateral |
|
Pia mater |
bundle |
|
eminence |
|
|
|
|
Lateral |
Basi- |
23 – 10 |
|
ventricle |
|
cranium |
|
|
|
|
Skeletogenous layer
Dural limiting layer
Cistern
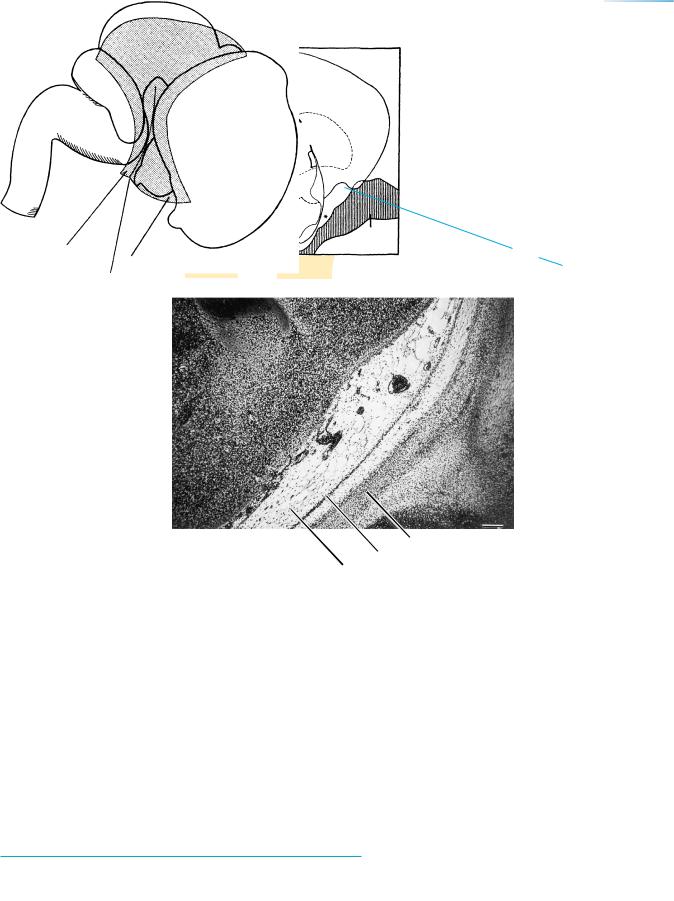
Figure 23–10. A coronal section showing a subarachnoid cistern (asterisk in key) situated between the olfactory bulb and the optic nerve. Most of the cisterns found in the adult are now present. Bar: 0.2 mm.
23 – 12
Caudolateral part
Rostrolateral part
Tentorial notch
Figure 23–11. Right lateral reconstruction to show (stippled) the tentorium cerebelli. The medial part of the tentorium still extends from the sellar region to the mamillary area, but it is becoming thinner. It separates incompletely two future subarachnoid areas: (1) that of the telencephalon and diencephalon, and (2) that of the rhombencephalon, including the cerebellum. In this reconstruction only the lateral parts are represented: the rostrolateral and the caudolateral, joined at the future tentorial notch, which is continuous with the medial part. The development of the tentorium during the fetal period is complicated, but has been well schematized by Markowski (1922) and measured by Klintworth (1967).
|
THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
233 |
B |
Permanent |
|
|
layers |
|
1. Pigment layer
3. ELM
Developmental
layers
External "neuroblastic"
Transient
fiber
Internal "neuroblastic"
9. N. fiber layer
10. ILM
C
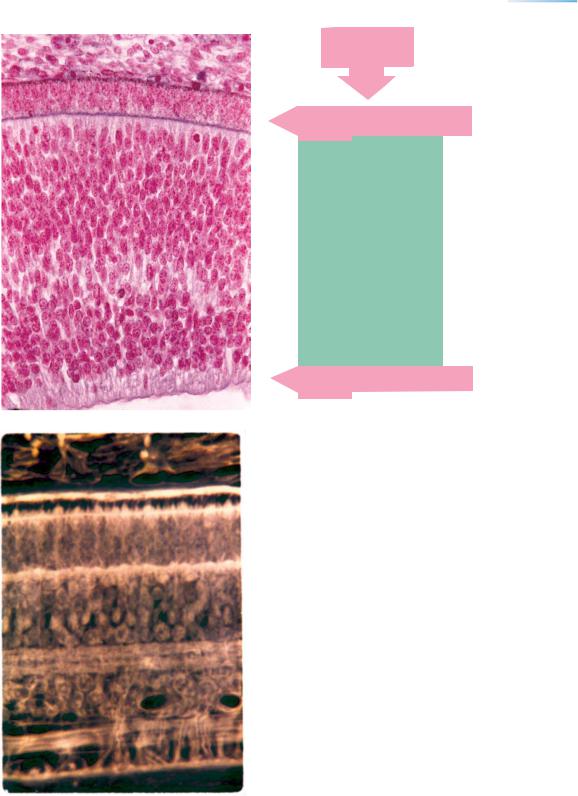
Figure 23–13. (Continued ) (B) The retina at 7 weeks (stage 20). Of the 10 layers that are characteristic later, layers 1, 3, 9, and 10 are present here. ×400.
Of the direct derivatives of the neuroepithelium of the optic cup (Figs. 14–6 and, 15–8), the external “neuroblastic” layer will give rise to layers 4, 5, and 6 (external nuclear, external plexiform, and internal nuclear) and the transient fiber zone to layer 7 (internal plexiform), both “neuroblastic” layers will contribute to layer 8 (ganglion cells).
(C) The retina at the end of trimester 2 (25 weeks). All 10 layers have been identifiable since approximately 15 weeks. For details of retinal development see Rhodes (1979).
ELM, ILM, external and internal limiting membranes.
234 |
C h a p t e r 2 3 : THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
|
|
Hipp. |
|
|
Nucleus |
|
|
accumbens |
|
|
V3 |
|
Basi- |
Chiasma |
|
sphenoid |
|
|
|
Basi- |
|
|
occipital |
23 – 15 14 |
|
& dens |
|
|
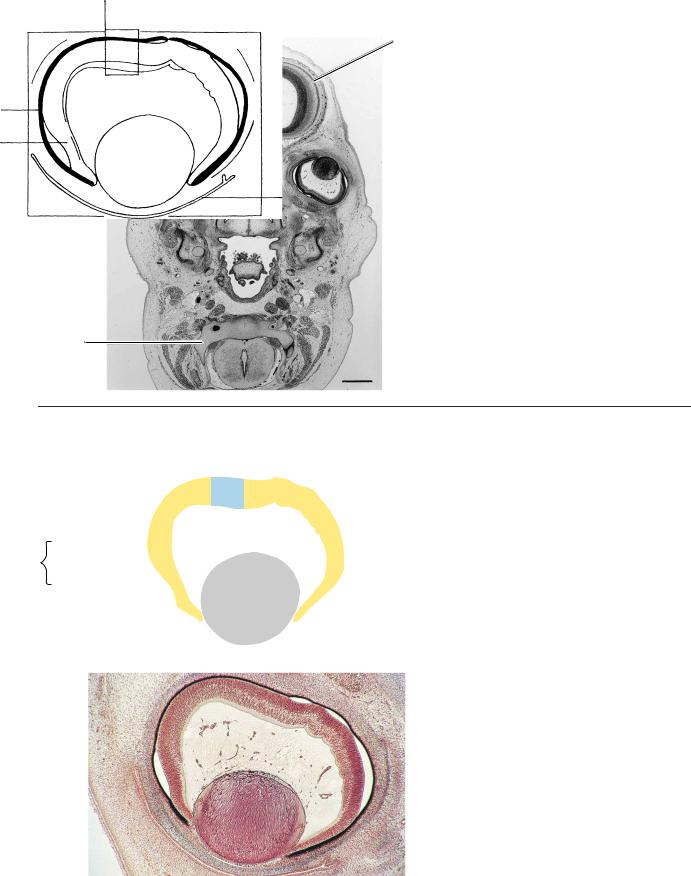
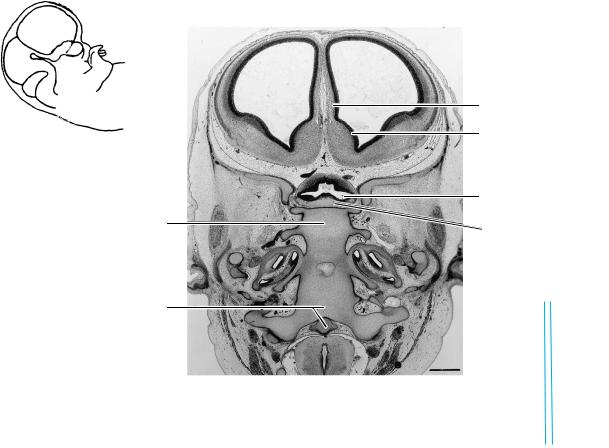
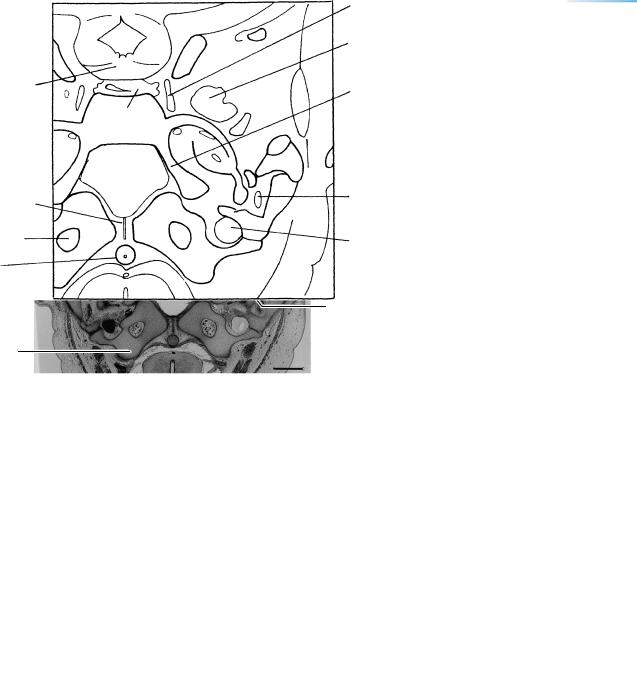
Figure 23–14. A section along the base of the skull. The internal ear is shown on each side. A small, basal part of the diencephalon can be seen at the level of the tuberculum sellae. The optic chiasma and the third ventricle are visible. The mesenchyme surrounding the diencephalon and telencephalon basally is the meshwork of a basal cistern. The cerebral hemispheres contain the septal area with the nucleus accumbens, the hippocampus, and the lateral ventricular eminence, which last is the primordium of the caudate nucleus. Ontogenetically, the nucleus accumbens is more closely related to the corpus striatum than to the septal nuclei. In addition, its origin is possibly associated with the medial ventricular eminence (Sidman and Rakic, 1982).

THE BRAIN AT THE END OF THE EMBRYONIC PERIOD |
237 |
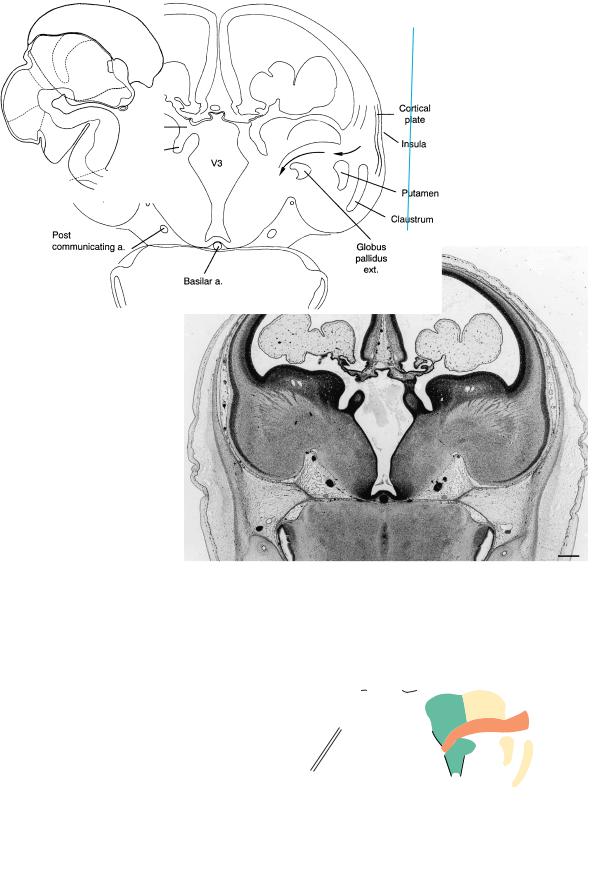
Figure 23–16. A section through the insular region and the left interventricular foramen. The cortical plate, which first appeared at stage 21 in the insular region, is thickest there, partly because of the increase of the intercellular spaces. The internal capsule, putamen, and globus pallidus externus are visible. The gray band lateral to the putamen is the claustrum. The choroid plexus at this time has become lobular, the epithelium is low prismatic, and the gelatinous stroma resembles that in the umbilical cord (Ariens¨ Kappers, 1966). The insula becomes buried during the fetal period and will possess extensive connections, as well as numerous functions (Augustine, 1996).
In the key drawing, arrows indicate the course of the internal capsule. Its fibers, mostly thalamocortical, become dispersed in the intermediate layer and the subplate of the neopallium. Some fibers may be corticothalamic, although it is doubtful that they reach the thalamus already.
The developing corpus striatum is composed of several neurochemically defined populations, and neuropeptide genes are expressed during trimester 2 (Brana et al., 1995). Moreover, the subependymal germinal layer is voluminous at this time, whereas the matrix disappears near birth. Neuropeptide gene expression is already under dopamine control through innervation from the substantia nigra (Freeman et al., 1995). The density of dopaminergic innervation is greatest caudal to the genu of the corpus callosum (Verney et al., 1993). Most, if not all, striatal neurons bear dopamine receptors (Brana et al., 1996). In addition to the nigrostriatal influence, corticostriatal inputs are believed to be important.
Figure 23–17. The sulcus terminalis is the site of the future thalamostriate vein and is parallel to the stria terminalis, which is recognizable already at stage 22 (Fig. 22–6B). The order of structures, from medial to lateral, is (1) hippocampus (still prominent rostrally), (2) choroid fissure and plexus, (3) thalamus and lamina affixa, (4) sulcus terminalis, and (5) caudate nucleus. Arrows indicate the internal capsule (in orange-brown).
Later in prenatal life the corpus callosum and the fornix become interposed between the rostral remains of the hippocampus and the choroid fissure. Then the order of structures is similar to that in the adult, as shown in the lower drawing: (1) indusium griseum and longitudinal striae (rostral remains of the hippocampus), (1a) corpus callosum and fornix, (2) choroid fissure and plexus, (3) thalamus and lamina affixa,
(4) thalamostriate vein and stria terminalis (in the sulcus terminalis), and (5) caudate nucleus.
The internal capsule (arrows) passes first between two telencephalic structures (finer stippling), the caudate nucleus and the putamen, and then between two diencephalic structures (coarser stippling), the thalamus and the globus pallidus.